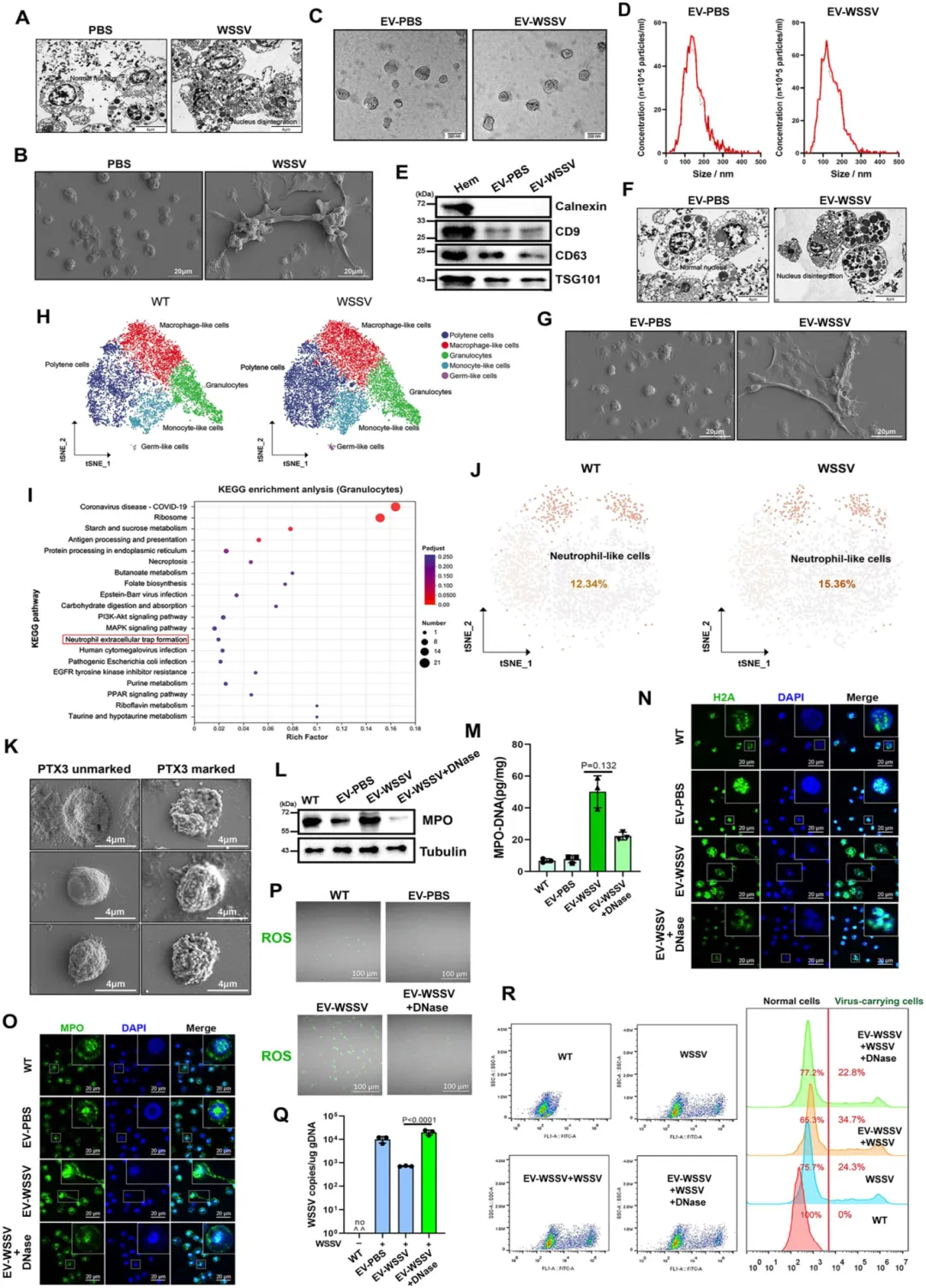
EVs 的核心功能:诱导类 NETosis 并抑制病毒
从 WSSV 感染泥蟹的血淋巴中分离出杯状 EVs(直径 100-200 nm),表达 EVs 标志物(CD9、CD63、TSG101)。
EV-WSSV(感染组 EVs)可单独诱导无病毒泥蟹的血细胞出现核解体、胞外捕获网结构(类 NETosis 典型特征),而 EV-PBS(对照组)无此效果。
EV-WSSV 处理后,血细胞中 MPO 蛋白、MPO-DNA 复合物、活性氧(ROS)水平显著升高,病毒拷贝数减少,病毒携带细胞比例下降,泥蟹存活率提高。
图1.EVs感染期间通过诱导类NETosis反应抑制病毒复制
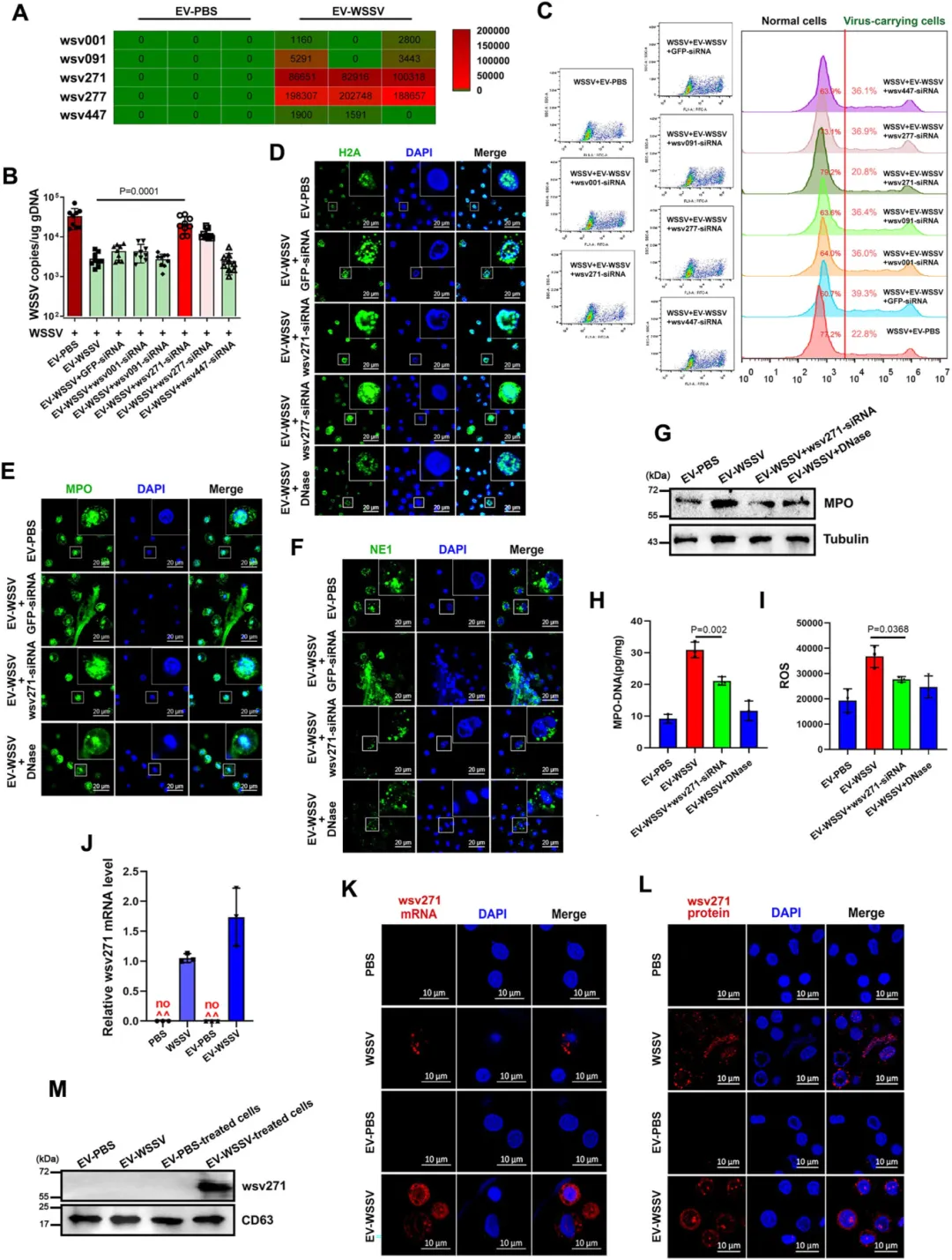
关键调控因子:wsv271 mRNA 是核心介导者
转录组测序发现,EVs 中仅病毒 mRNA wsv271、wsv277 与 EVs 的抗病毒功能相关,其中 wsv271 是必需因子。
EVs 可将 wsv271 mRNA 传递至受体血细胞,且该 mRNA 能在细胞内翻译为病毒蛋白(EVs 本身不携带 wsv271 蛋白)。
沉默 wsv271 后,类 NETosis 相关指标(如胞外捕获网、ROS 水平)显著减弱,病毒抑制效果消失,病毒携带细胞比例回升。
图2.EVs通过包裹wsv271 mRNA调控类NETosis反应与病毒复制
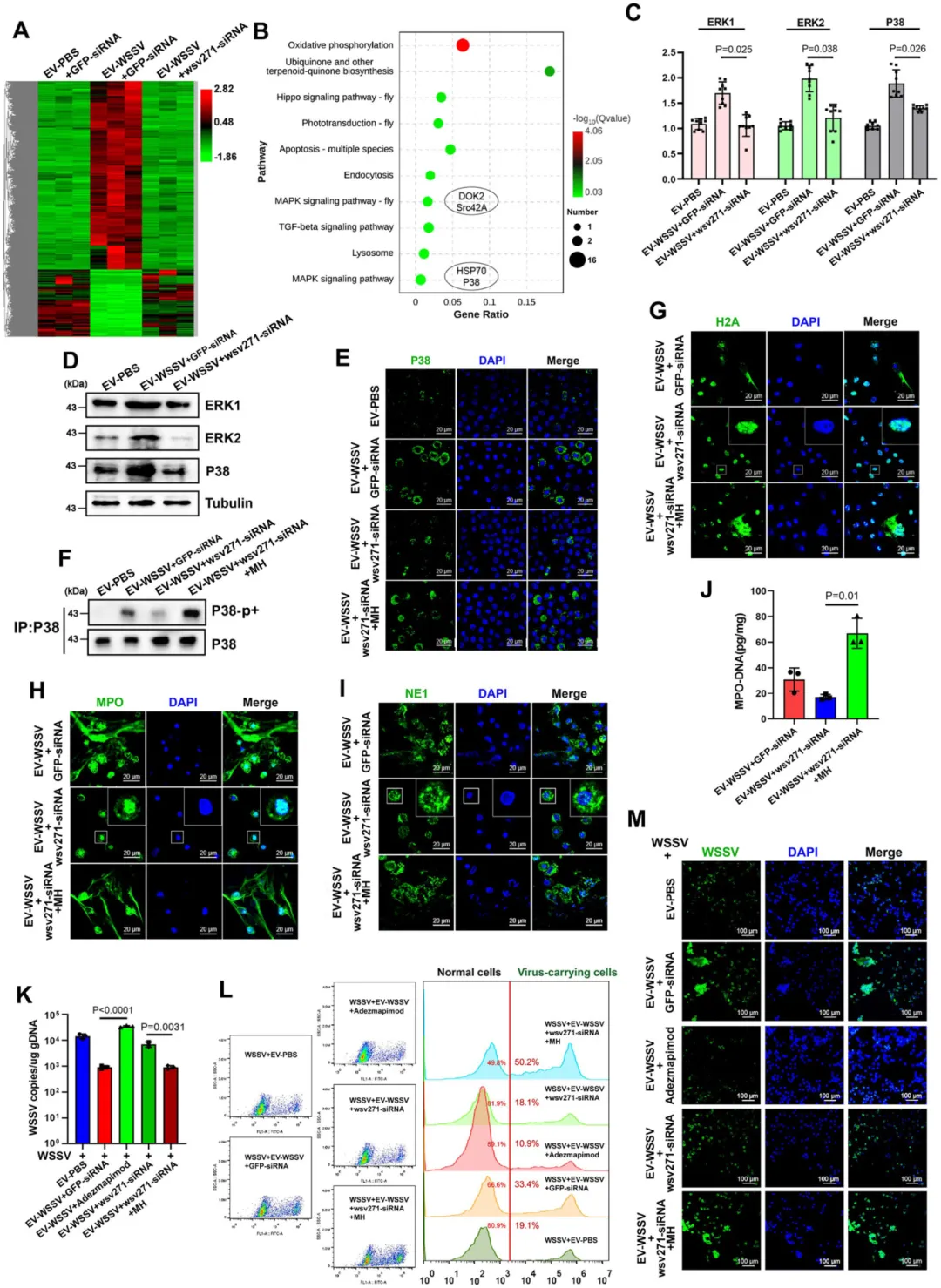
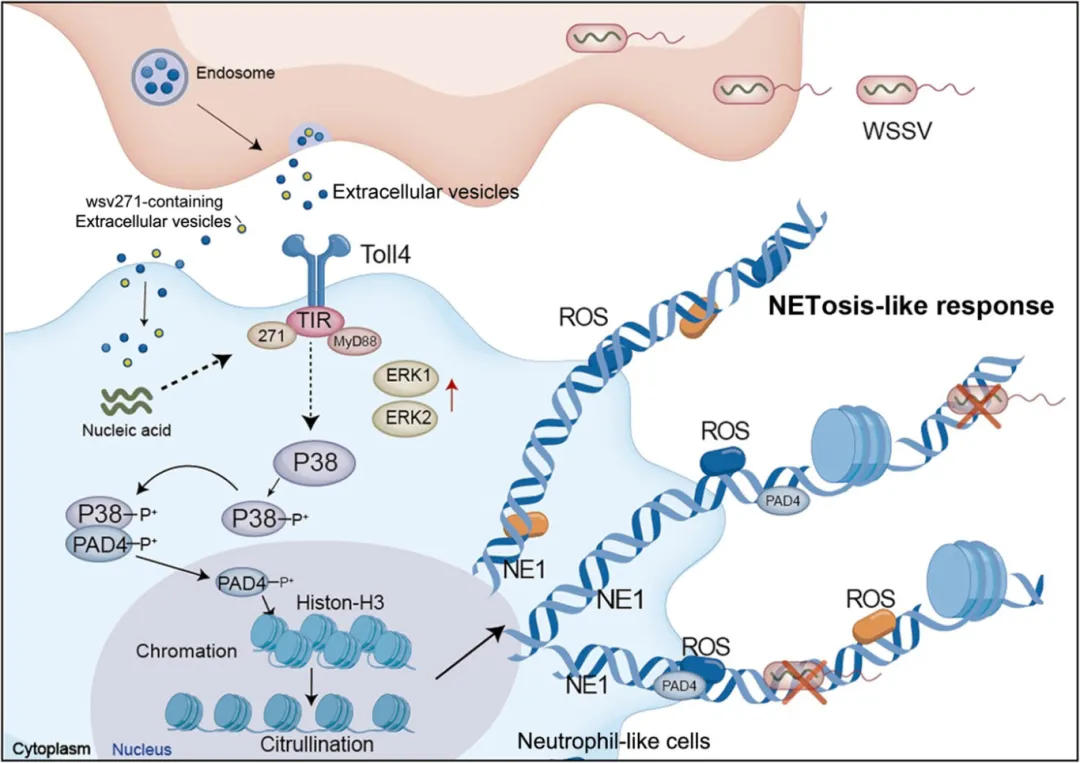
信号通路激活:wsv271-Toll4-MyD88-P38-MAPK 轴
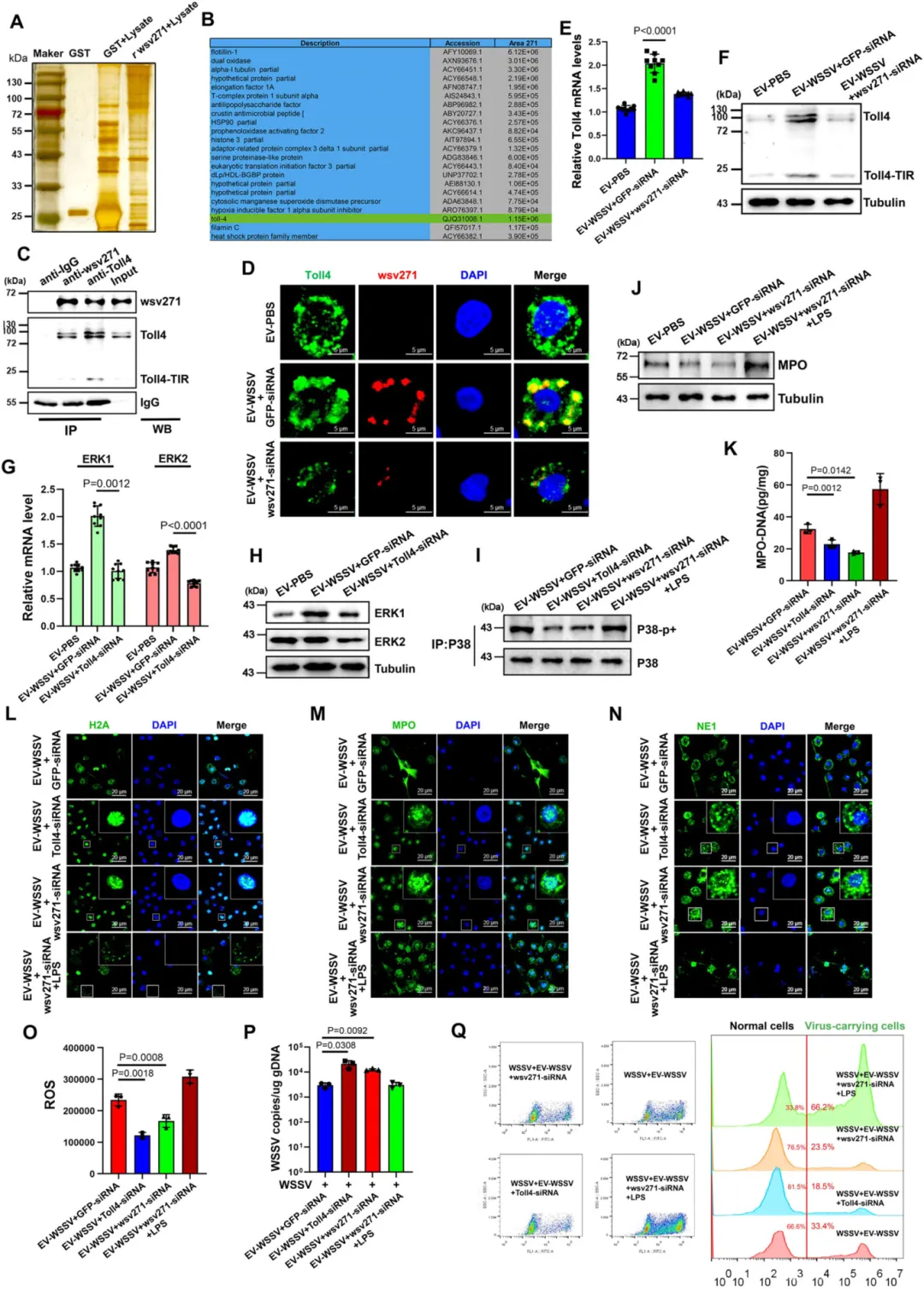
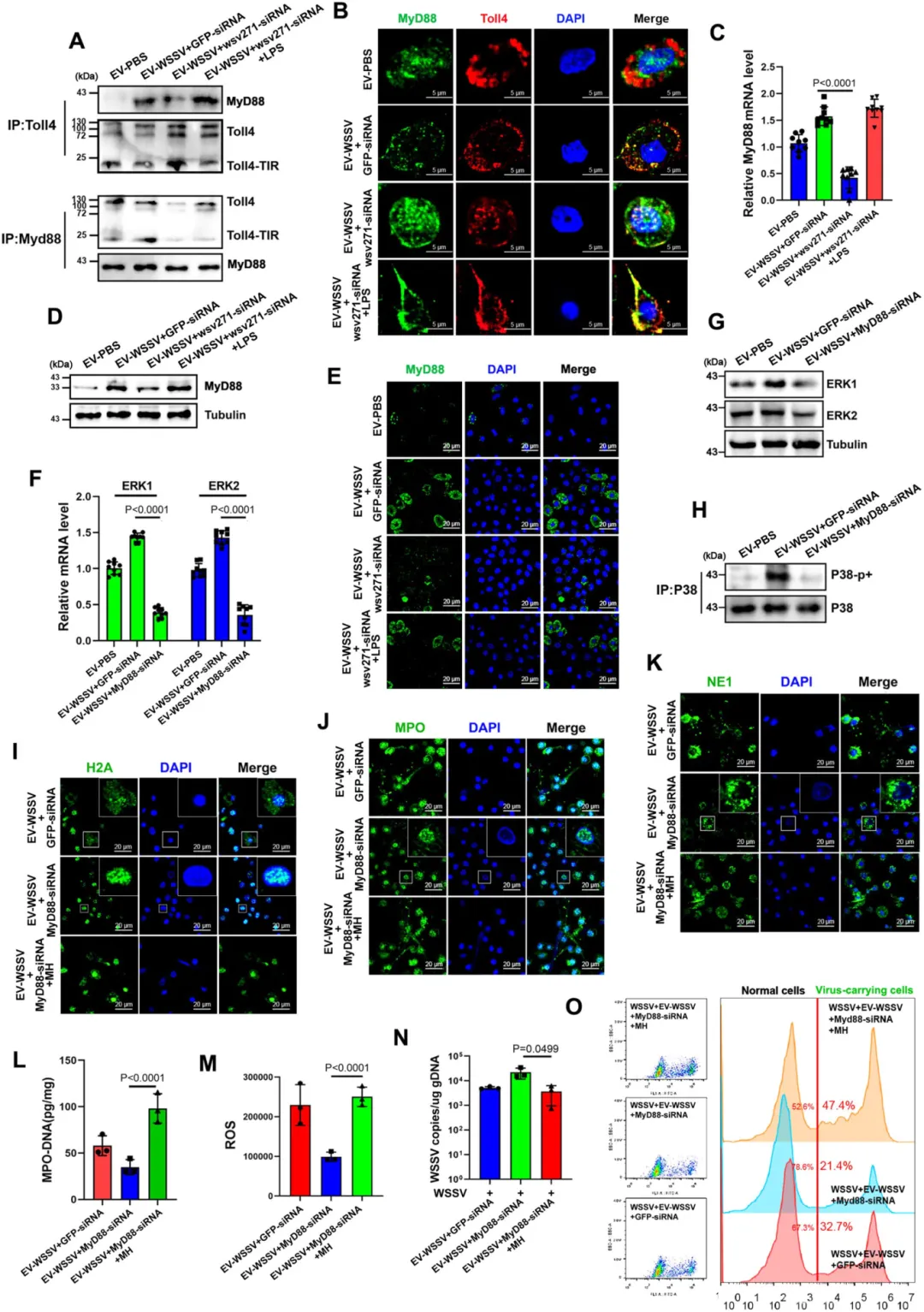
GST-pull down 和免疫沉淀验证:wsv271 蛋白与 Toll4 的 TIR 结构域直接相互作用。
该相互作用可招募 MyD88,进而激活 P38-MAPK 通路(ERK1、ERK2、P38 的 mRNA 和蛋白水平升高,P38 磷酸化增强)。
沉默 Toll4 或 MyD88,P38-MAPK 通路激活受阻,类 NETosis 反应减弱,病毒抑制效果下降;加入通路激活剂(LPS、MH)可恢复该反应。
图3.EVs包裹的wsv271激活P38-MAPK通路调控类NETosis反应与病毒复制
图4.wsv271通过与Toll4相互作用激活P38-MAPK通路
图5.Toll4通过招募MyD88激活wsv271依赖的P38-MAPK通路
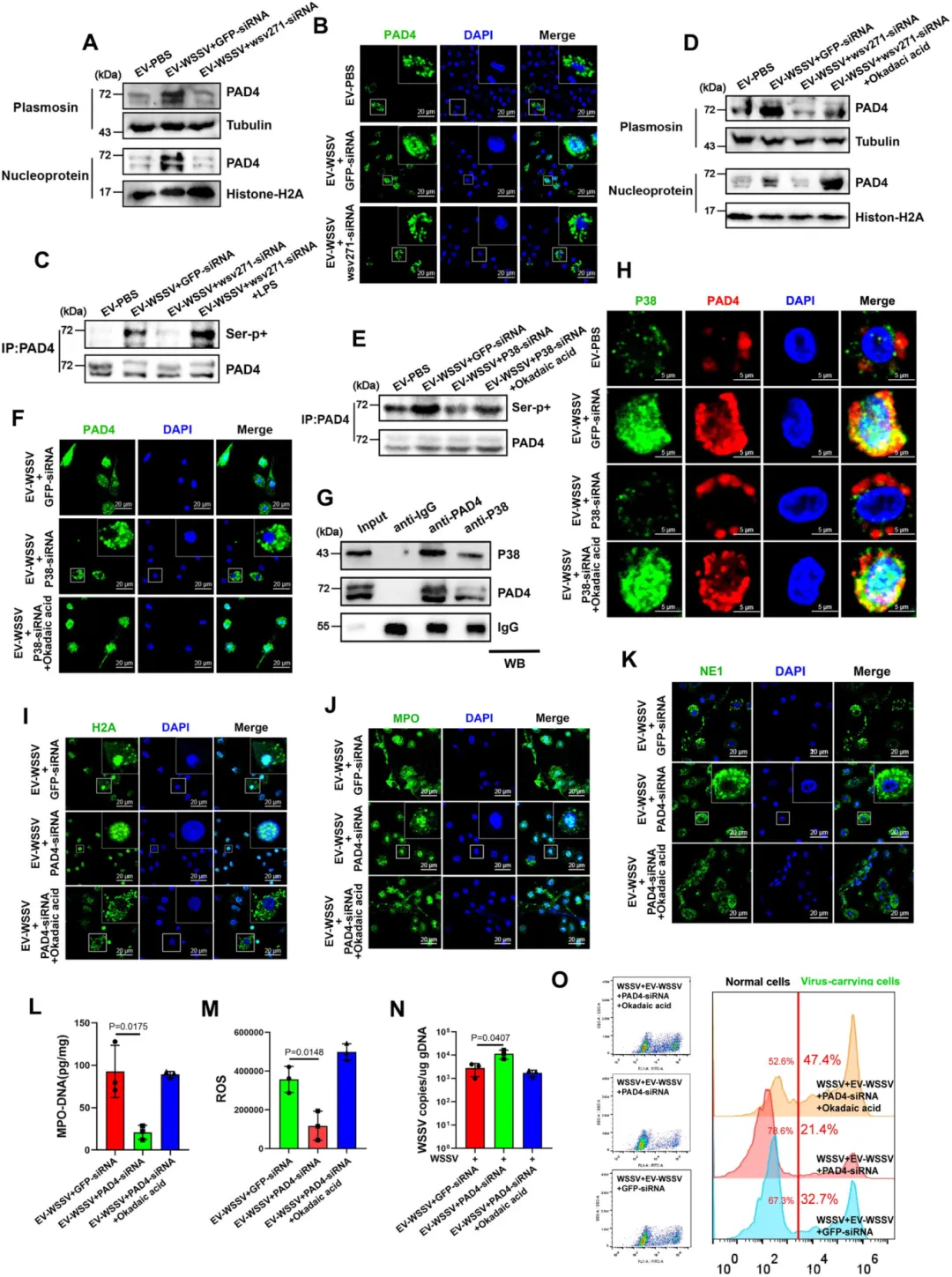
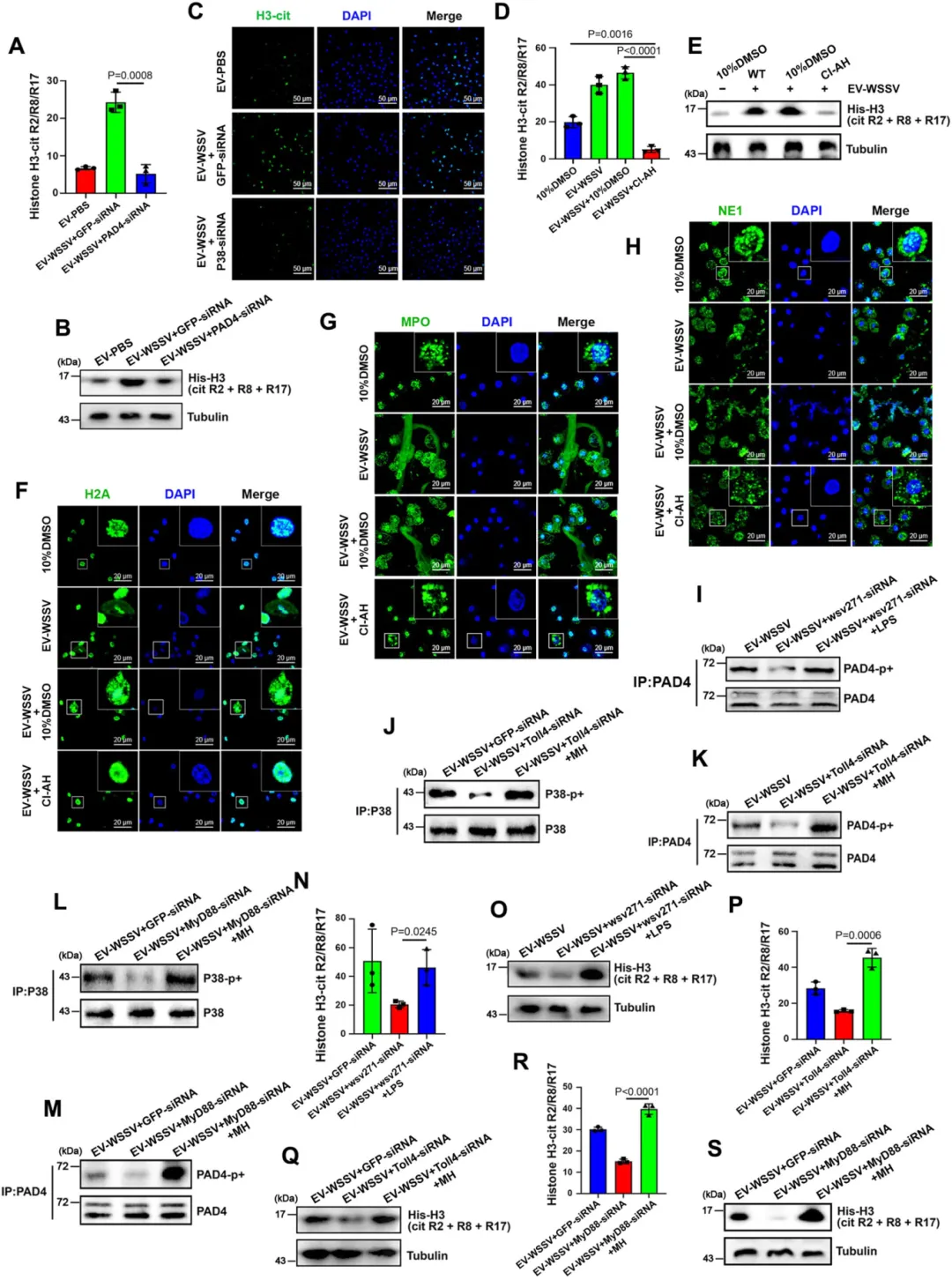
P38-MAPK 通路激活后,P38 可直接磷酸化 PAD4 蛋白,并促进其核转位(核内 PAD4 含量显著增加)。
核内磷酸化的 PAD4 催化组蛋白 H3 瓜氨酸化,这是类 NETosis 启动的关键开关(抑制 PAD4 或组蛋白瓜氨酸化,胞外捕获网无法形成)。
图6.P38介导的PAD4磷酸化与核转位是类NETosis反应激活的必需条件
图7.磷酸化PAD4催化组蛋白H3瓜氨酸化以激活类NETosis反应
图8.EVs来源病毒核酸诱导类NETosis反应的机制示意图