SM-1-TA和SM-2-TA:来自Streptomyces avermitilis的两种单宁酶同工酶的异源表达、酶学/结构表征及功能互补性
近日,南昌大学食品学院食品营养与健康创新团队的硕士研究生曾欢、邓泽元教授、张鹏副教授等人在国际期刊Journal of Food Science发表了题为
“SM-1-TA and SM-2-TA: Heterologous Expression, Enzymatic/Structural Characterization, and Functional Complementarity of Two Tannase Isoenzymes From Streptomyces avermitilis”
的文章。单宁酶是一种在食品工业等领域具有广泛应用潜力的工业酶。本研究从阿维链霉菌中鉴定出两种单宁酶同工酶SM-1-TA和SM-2-TA,并对其生物信息学和酶学性质进行了比较分析。SM-1-TA和SM-2-TA的最适温度分别为35°C和45°C,最适pH分别为7.0和8.0。值得注意的是,SM-2-TA表现出高热稳定性,在40°C下孵育2小时后仍保留40.03%的残余活性。大多数金属离子和化学试剂对SM-1-TA和SM-2-TA的催化活性均有抑制作用。同时,与SM-1-TA相比,SM-2-TA对有机试剂表现出更高的耐受性。以没食子酸甲酯、没食子酸丙酯和单宁酸为底物时,SM-1-TA的kcat/Km分别为77.59、63.53和198.31 s⁻¹ mM⁻¹,而SM-2-TA的kcat/Km分别为44.65、23.43和473.67 s⁻¹ mM⁻¹。分子动力学模拟表明,SM-1-TA较低的热稳定性可能源于其较高的整体柔韧性和残基区域100-150内显著的局部波动。本研究中观察到的SM-1-TA和SM-2-TA之间的功能互补性突显了同工酶在微生物适应不同环境中的生态学意义。这些发现不仅为理解同工酶的功能分化提供了见解,也为未来发现和工程化具有增强性能的单宁酶奠定了基础。
(原文链接)https://ift.onlinelibrary.wiley.com/doi/10.1111/1750-3841.71040
单宁酶(单宁酰基水解酶,E.C. 3.1.1.20)催化水解单宁中的酯键和缩酚酸键(酚酸与羟基之间形成的酯),释放没食子酰基酯、葡萄糖和没食子酸。这种催化功能使单宁酶成为一种重要的工业酶,并已广泛应用于食品、饮料、制药和动物饲料等多个领域。单宁酶在生物分类中分布广泛,可从植物、动物以及微生物系统中分离得到。特别是源自微生物的单宁酶,凭借其在生物多样性和高产率方面的优势,已取代了传统的植物和动物来源,使得微生物发酵成为主要的工业生产模式。
单宁是植物次生代谢产物,在防御生物胁迫中发挥作用。单宁被认为是一种抗营养物质,能与蛋白质等大分子形成水不溶性复合物,也是茶饮料涩味的主要来源。因此,在各个行业中,基于单宁酶的生物降解方法因其环境友好性和生态友好性,已被广泛应用于解决由单宁引起的各种问题。例如,在食品工业中,Govindarajan等人发现,绿茶汤经单宁酶处理后,茶汤中茶乳酪沉淀量显著减少,同时茶汤香气更浓,口感更醇厚,涩味适中。在环境保护领域,Abirami等人利用单宁酶处理制革废水,旨在通过降低单宁酸含量来提高皮革厂废水处理站的处理效率。
在以往的研究中,单宁酶的克隆、异源表达和表征方面已取得重大进展。然而,与其他生物来源的单宁酶相比,微生物来源的单宁酶更稳定,已成为当前研究的重。目前,商业单宁酶主要通过真菌发酵获得。例如,Shao等人报道了从黑曲霉FJ0118中克隆并表征一种新型单宁酶。然而,与真菌来源的单宁酶相比,细菌来源的单宁酶由于其结构特征,更容易克隆、过表达和纯化。因此,细菌来源的单宁酶可能成为真菌来源单宁酶的理想替代品。Kanpiengjai等人从乳酸菌中鉴定出一个碱性单宁酶基因,并在大肠杆菌BL21(DE3)中表。尽管如此,对细菌单宁酶的研究仍然有限,只有少数单宁酶基因被克隆和重组表达。因此,发现能够高效水解天然底物的新型、高活性细菌单宁酶,为改善单宁的酶法水解提供了一种有前景且成本效益高的策略。
此外,同工酶(催化相同反应但结构或动力学特性不同的酶)的进化是微生物响应环境波动的一种适应性策略。在细菌中,这种功能不同的同工酶的发展使得在稳定条件下能够优化底物利用,同时在胁迫下保持催化韧性。例如,从轮枝菌属P9菌株中分离的两种冷适应单宁酶在宽温度范围内显著增强了单宁的生物降解,说明了同工酶分化如何扩展功能多样性。在之前的研究中,我们还从链霉菌属303MFCol5.2菌株中鉴定出两种具有不同酶学特性的单宁酶,这可能有助于提高宿主生物的环境适应性。
本研究从阿维链霉菌中成功分离出两种新型单宁酶,以扩大单宁酶的细菌来源,命名为SM-1-TA和SM-2-TA,并实现了它们在大肠杆菌BL21(DE3)中的表达。对重组SM-1-TA和SM-2-TA的生物信息学特征和生化性质进行了系统研究。此外,采用分子对接和分子动力学模拟初步阐明了这些酶的潜在催化机制。这项工作为单宁酶的未来研究和应用奠定了理论基础。
1)、SM-1-TA和SM-2-TA的生物信息学特征
新型单宁酶的发现有望拓宽其在食品工业及相关领域的应用。通过基因挖掘,在阿维链霉菌中鉴定出两个编码推定单宁酶的基因。相应的酶被命名为SM-1-TA(GenBank:BAC69254.1)和SM-2-TA(GenBank:WP_010983463.1)。基于从NCBI数据库检索到的开放阅读框,分析了SM-1-TA和SM-2-TA的生物信息学特征。
经SignalP 6.0进行信号肽预测和去除,SM-1-TA被鉴定为由507个氨基酸组成的酸性蛋白,理论等电点为5.12,分子量为53.41 kDa。它包含56个带负电的氨基酸(Asp + Glu)和42个带正电的氨基酸(Arg + Lys)。相比之下,SM-2-TA是由476个氨基酸组成的酸性蛋白,理论pI为5.61,Mw为50.54 kDa。它包含49个带负电的氨基酸(Asp + Glu)和41个带正电的氨基酸(Arg+Lys)。SM-1-TA和SM-2-TA的不稳定指数分别为22.63和21,均小于40,表明它们是稳定蛋白。二级结构预测表明,SM-1-TA由32.15%的α-螺旋、9.27%的延伸链、3.16%的β-转角和55.42%的无规则卷曲组成。SM-2-TA由33.61%的α-螺旋、13.24%的延伸链、1.89%的β-转角和51.26%的无规则卷曲组成。
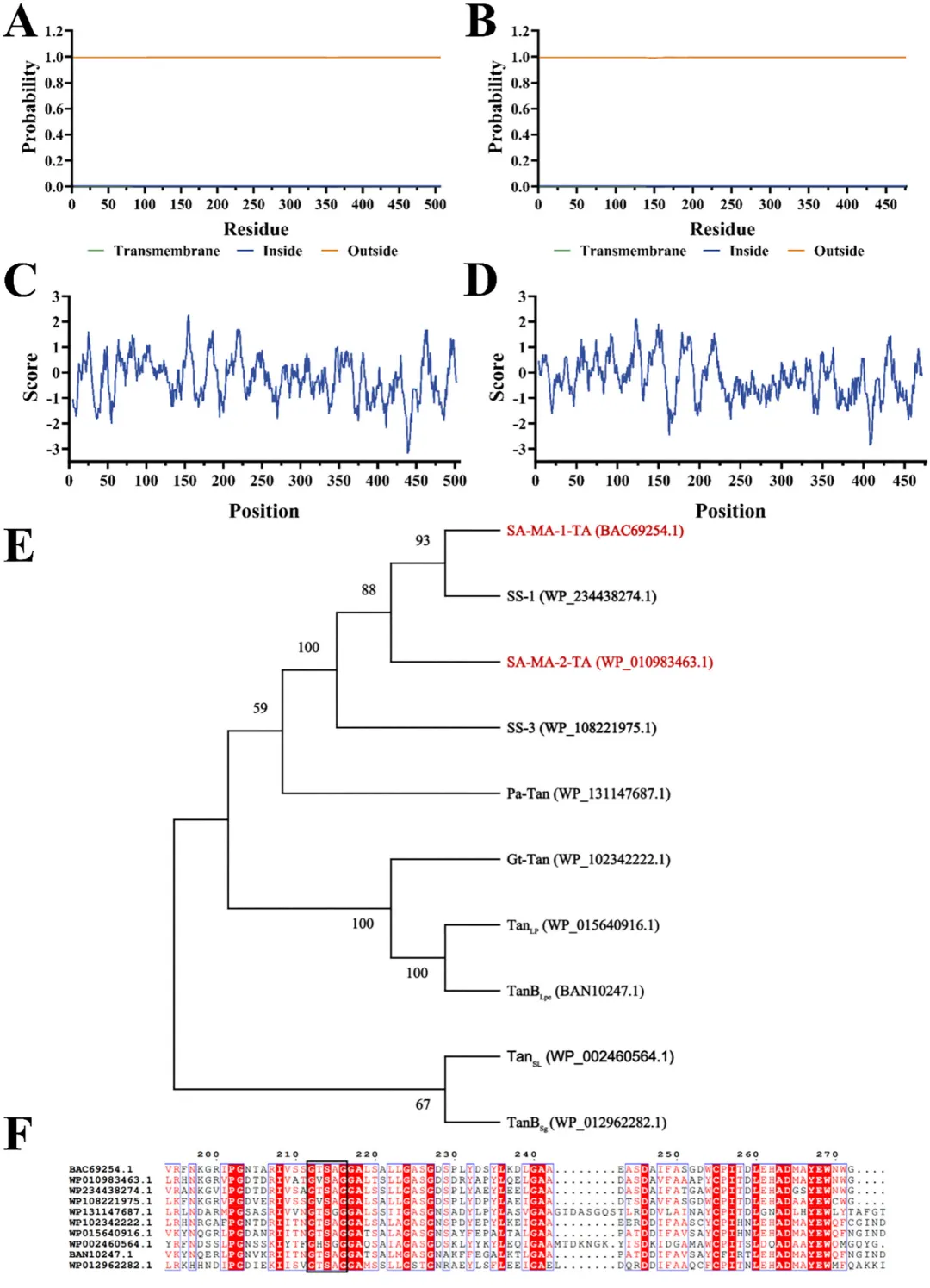
TMHMM分析的跨膜结构表明,SM-1-TA和SM-2-TA缺乏跨膜结构(图1A、B)。使用ProtScale预测了SM-1-TA和SM-2-TA蛋白的疏水性和亲水性。如图1C所示,SM-1-TA蛋白的第154位氨基酸表现出最高的疏水值2.233,而第438位氨基酸表现出最高的亲水值-3.156。相比之下,如图1D所示,SM-2-TA蛋白的第123位氨基酸表现出最高的疏水值2.111,而第408位氨基酸表现出最高的亲水值-2.8333。根据亲水性总平均值,SM-1-TA和SM-2-TA被归类为亲水性蛋。
为了阐明SM-1-TA和SM-2-TA的进化关系,使用MEGA 11.0软件构建了SM-1-TA和SM-2-TA的系统发育树。结果表明,单宁酶具有共同起源,但分化为两个不同的分支(图1E)。其中,SM-1-TA和SM-2-TA与SS-1处于同一子分支,它们与SS-1的相似性分别为68.88%和59.04%。将SM-1-TA和SM-2-TA的氨基酸序列与其他报道的单宁酶进行了比较。图1F显示,所有单宁酶都具有保守的五肽基序“Gly-X-Ser-X-Gly”。此外,对比对序列的分析鉴定出推定的催化三联体为SM-1-TA中的Ser214-Asp454-His486和SM-2-TA中的Ser182-Asp423-His455。
2)、SM-1-TA和SM-2-TA的分子结构与分子对接
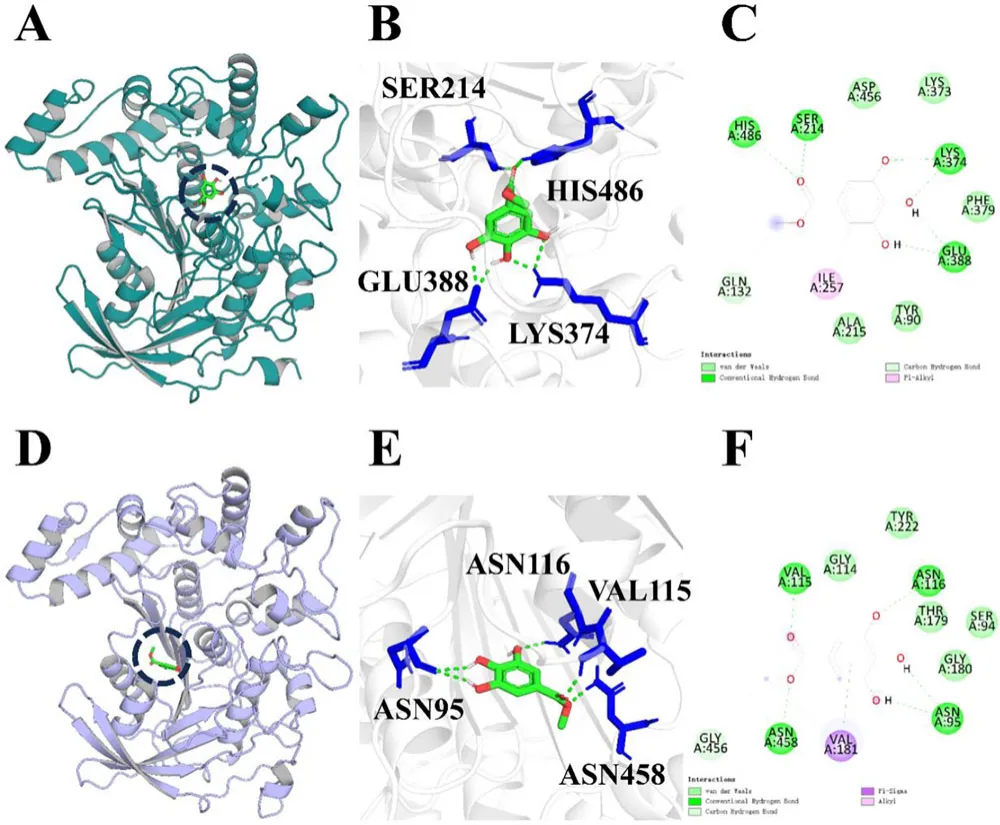
催化三联体对于单宁酶活性至关重要,这一点已在先前研究中确立。因此,采用分子对接进一步研究底物与单宁酶之间的相互作用。通过AlphaFold2构建了SM-1-TA和SM-2-TA的3D结构。使用两个独立的指标验证了模型质量。Ramachandran图分析证实,SM-1-TA和SM-2-TA分别有99.8%和99.5%的残基位于最适允许区。此外,Verify 3D评估显示,各自结构中有92.5%和88.24%的残基的平均3D-1D得分≥ 0.1,超过了可靠模型80%的阈值。上述结果表明预测的蛋白质模型具有很高的可信度。
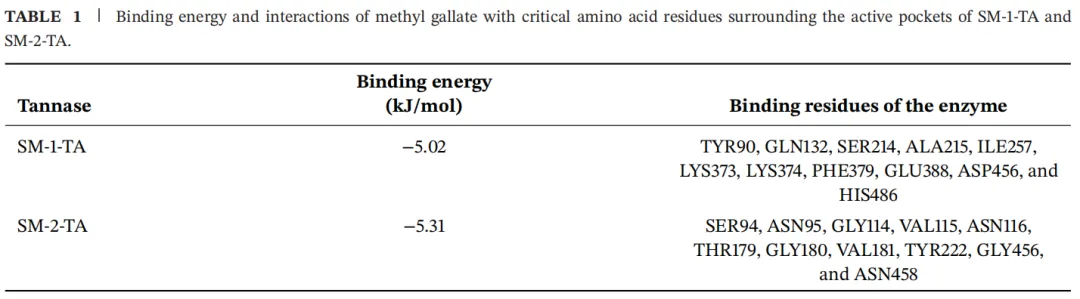
为了进一步研究底物与SM-1-TA和SM-2-TA之间的相互作用,使用没食子酸甲酯作为底物进行分子对接(图2)。氢键在促进酶-底物相互作用中起着关键作用。其中,没食子酸甲酯与SM-1-TA中的4个氨基酸残基形成了6个常规氢键,包括SER214(1.87 Å)、LYS374(1.86 Å, 2.22 Å)、GLU164(1.95 Å, 1.95 Å)和HIS486(2.56 Å)。此外,没食子酸甲酯通过范德华力、碳氢键和Pi-烷基与7个残基相互作用,包括TYR90、GLN132、ALA215、ILE257、LYS373、PHE379和ASP456。同时,没食子酸甲酯与SM-2-TA中的4个氨基酸残基形成了5个常规氢键,包括ASN95(2.16 Å, 2.19 Å)、VAL115(1.89 Å)、ASN116(2.14 Å)和ASN458(1.95 Å)。此外,没食子酸甲酯通过范德华力、碳氢键、Pi-Sigma和烷基与7个氨基酸残基相互作用,包括SER94、GLY114、THR179、GLY180、VAL181、TYR222和GLY456。
分子对接的结合能是表征配体与受体之间热力学稳定性的重要指标。更负的结合能值表示配体与受体之间的结合亲和力更强,从而可以预测结合强度和潜在的催化效率。没食子酸甲酯与SM-1-TA和SM-2-TA的结合能分别为-5.02和-5.31 kJ/mol,表明它们之间形成了稳定的复合物(表1)。这表明配体和受体之间存在很强的亲和力。总之,SM-1-TA和SM-2-TA通过一系列分子间作用力与没食子酸甲酯形成复合物,从而促进没食子酸甲酯的水解。
3)、SM-1-TA和SM-2-TA的表达与生化表
3.1)、SM-1-TA和SM-2-TA的表达
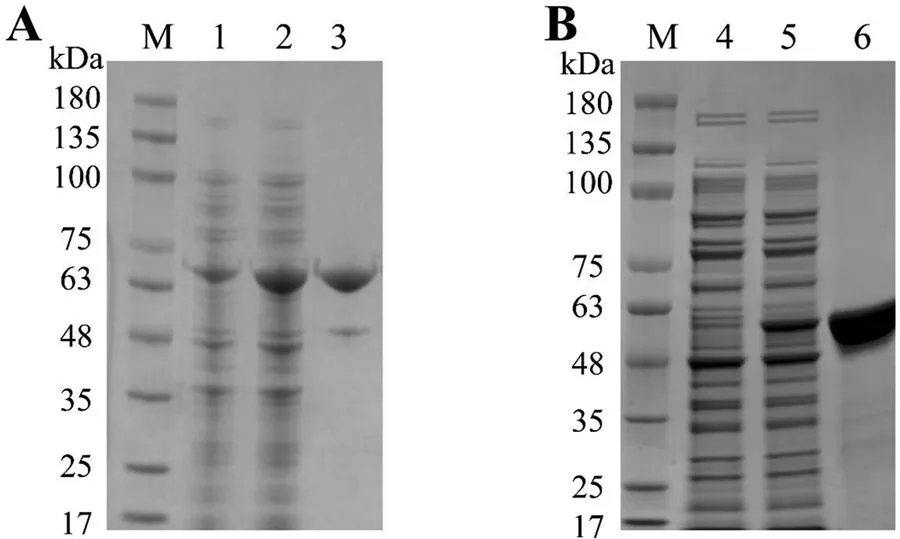
SM-1-TA和SM-2-TA蛋白通过IPTG诱导表达,然后通过镍柱层析纯化。SDS-PAGE分析显示,SM-1-TA和SM-2-TA在染色后呈现单一且显著的条带(图2)。观察到的分子量与理论预测相符。源自细菌的单宁酶分子量通常在31至90 kDa之间。结合SDS-PAGE结果,SM-1-TA和SM-2-TA均被确认具有细菌单宁酶特征的分子量(图3)。
3.2)、pH和温度对活性的影响
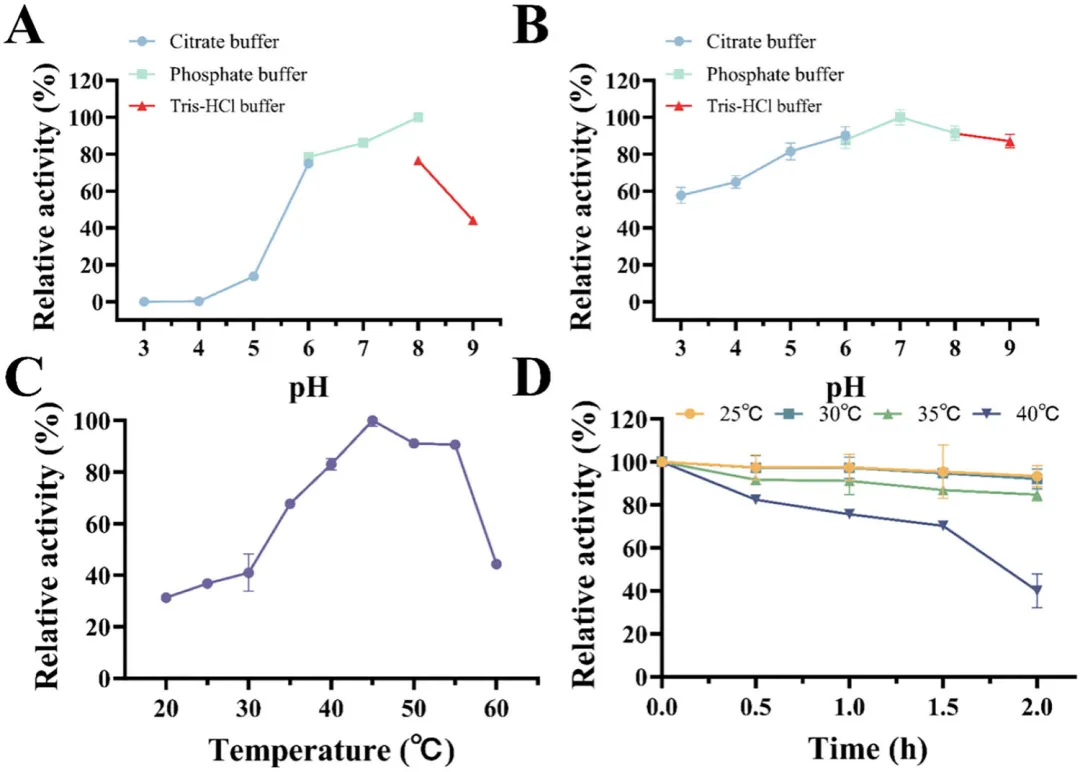
通常,pH和温度通过显著改变催化效率来关键影响酶的催化活性。细菌单宁酶通常在pH 5.0–7.0范围内表现出最佳活性。因此,在pH 3.0–9.0的范围内评估了SM-1-TA和SM-2-TA的活性。SM-1-TA和SM-2-TA的最适pH值分别为7.0和8.0(磷酸盐缓冲液)(图4A和5A)。两种酶在pH 6.0–8.0范围内均保留超过60%的活性,但在此范围之外,尤其是在pH低于5.0时,表现出显著的活性损失。同时,在不同pH条件下孵育24小时后,SM-1-TA和SM-2-TA在pH 5.0至8.0范围内均能维持超过60%的活性(图4B和5B)。值得注意的是,SM-2-TA在pH 3.0至5.0范围内能维持超过40%的活性,表明其具有一定的工业应用潜力。缓冲液性质(类型/浓度)可能调节酶的稳定性,需要进行系统研究以阐明这些复杂的相互作。
除pH外,温度也关键影响酶的活性。在20–60°C的温度梯度下评估了SM-1-TA和SM-2-TA的活性(图4C和5C)。SM-1-TA和SM-2-TA的最适温度分别为35°C和45°C。这些值与大多数细菌单宁酶报道的最适温度范围30–50°C一致。同时,两种酶在达到各自最适温度前,催化活性随温度升高而增加,超过最适温度后则观察到活性迅速下降。值得注意的是,SM-1-TA和SM-2-TA在35–50°C温度范围内能保持较高的酯酶活性,其相对酶活性均超过80%。在55°C时观察到显著差异,SM-2-TA保持90.67%的相对活性,显著高于SM-1-TA保留的30%活性。
热稳定性是工业酶应用的关键参数。如图4D和5D所示,SM-1-TA和SM-2-TA的热稳定性表明它们在35°C以下具有良好的稳定性。孵育2小时后,相对酶活性仍能保持在80%以上。在40°C孵育2小时后,SM-1-TA仅保留11.05%的残余活性,而SM-2-TA保留40.03%的残余活性。这种行为与其他细菌单宁酶的报道一致。例如,Wu等人报道的单宁酶Gt-Tan,在40°C孵育48小时后,仅保留54.7%的残余活性,而在50°C孵育6小时后,其活性完全丧失。总的来说,SM-2-TA更宽的pH耐受性和优异的热稳定性凸显了其在食品工业中应用的巨大潜力。
3.3)、金属离子、表面活性剂、抑制剂和有机试剂对活性的影响
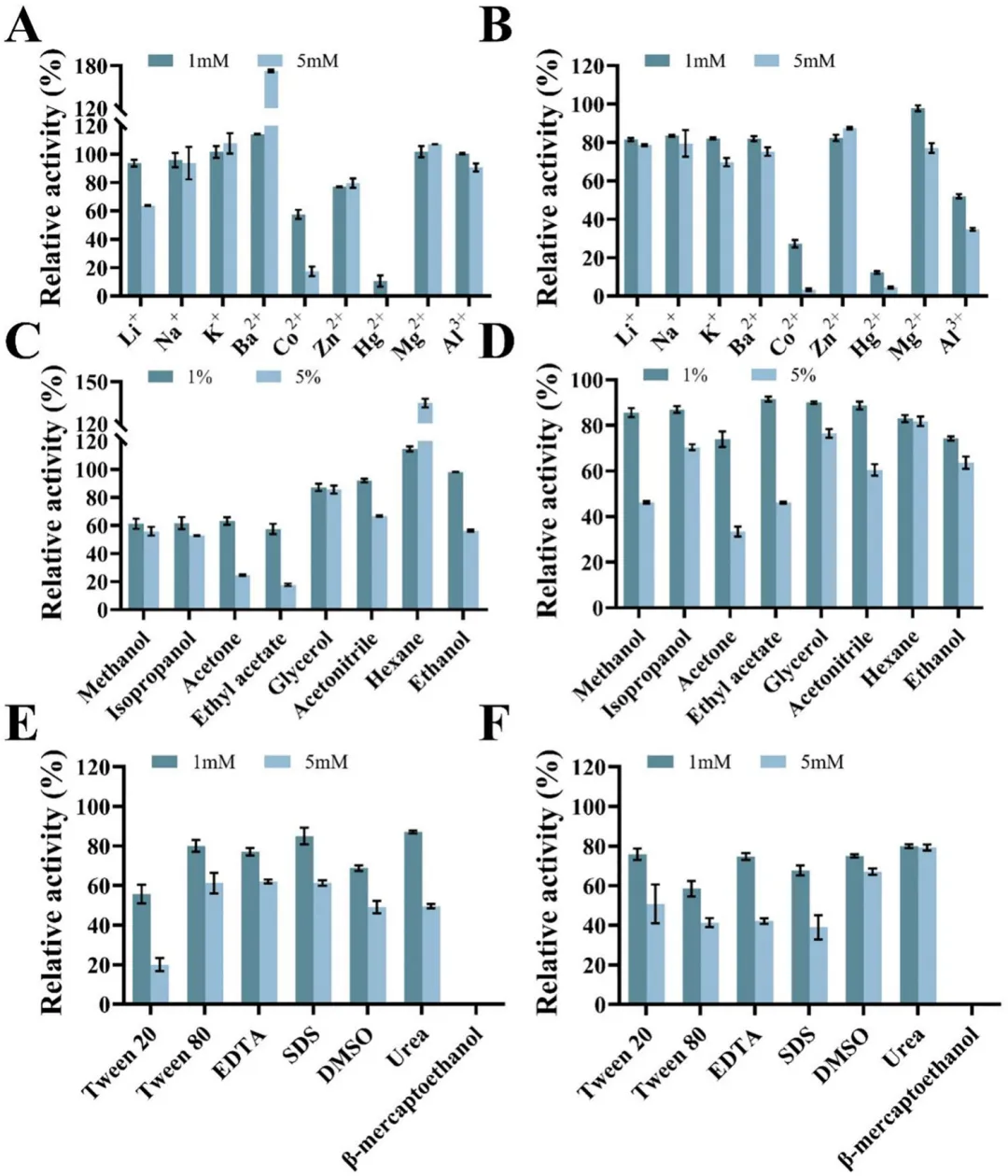
各种金属离子对SM-1-TA和SM-2-TA活性的影响如图6A和6B所示。其中,Co²⁺和Hg²⁺对两种酶均有强烈抑制作用。值得注意的是,在高浓度下,Hg²⁺的存在极大地抑制了SM-1-TA和SM-2-TA的活性,残余活性分别仅为0%和4.52%。这可能是由于金属离子与酶活性位点内的关键残基(如巯基、色氨酸或羧基)形成复合物,从而在空间上阻碍底物的结合。在高浓度Ba²⁺存在下,SM-1-TA的活性提高了72.6%,表明Ba²⁺可能与SM-1-TA底物结合口袋内的极性残基形成相互作用,从而稳定酶的催化构象并提高催化效率。
各种表面活性剂、抑制剂和有机试剂对SM-1-TA和SM-2-TA酶活性的影响如图6C-6F所示。大多数测试的试剂对两种酶都有不同程度的抑制。值得注意的是,β-巯基乙醇完全抑制了SM-1-TA和SM-2-TA的酶活性,这可能是由于破坏了其活性位点的分子间作用力,从而影响酶活性。同时,己烷增强了SM-1-TA的酶活性,在1%和5%的浓度下,其活性分别提高了14.56%和34.77%。相比之下,所有其他有机试剂对SM-1-TA和SM-2-TA的酶活性均有不同程度的抑制,且抑制效应随浓度增加而增强。这可能是由于有机试剂减少了酶微环境中必要的水分含量,导致蛋白质变性。总之,SM-1-TA和SM-2-TA在低浓度下对多种化学试剂均表现出较强的耐受性,显示出相当大的工业应用潜力。
单宁酶SM-1-TA和SM-2-TA的储存稳定性如图S5所示。在4°C储存条件下,两种酶都保持了相对较高的稳定性。储存10天后,SM-1-TA保留了79.53%的初始酶活性,而SM-2-TA保留了高达91.51%的初始酶活性。与SM-2-TA相比,SM-1-TA表现出较低的储存稳定性,这可能归因于其较弱的热稳定性。在长期储存过程中,其蛋白质构象容易发生结构松弛和扰动,导致催化活性降低。
3.4)、SM-1-TA和SM-2-TA的动力学参数
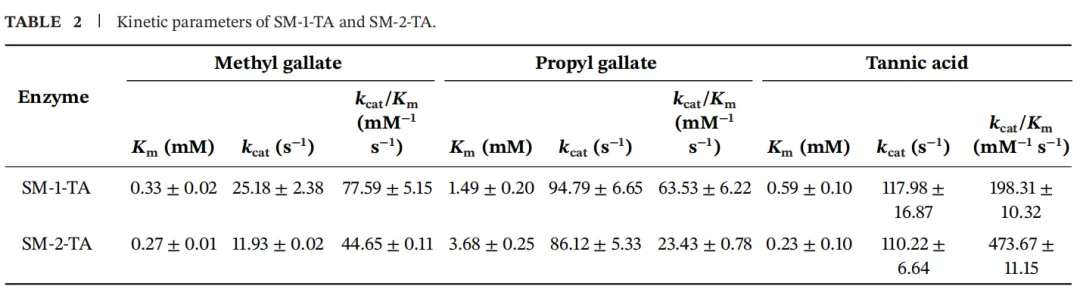
在本研究中,使用三种底物(没食子酸甲酯、没食子酸丙酯和单宁酸)测定了SM-1-TA和SM-2-TA的动力学参数。Km、kcat和kcat/Km的值列于表2。Km值是反映酶与其底物之间结合亲和力的关键指标,Km值越低表示亲和力越高。当使用没食子酸甲酯作为底物时,SM-2-TA和SM-1-TA的Km值分别为0.33 ± 0.02 mM和0.27 ± 0.01 mM,表明SM-2-TA对没食子酸甲酯具有更高的亲和力。这一观察结果与分子对接结果一致。这些同工酶不同的催化特性可能通过拓宽其底物利用谱,为生产生物提供适应性优势。此外,基于kcat/Km,在测试的三种底物中,SM-1-TA和SM-2-TA均对单宁酸表现出最高的催化效率,这与先前报道的其他单宁酶的动力学参数一致。
4)、SM-1-TA和SM-2-TA的分子动力学模拟
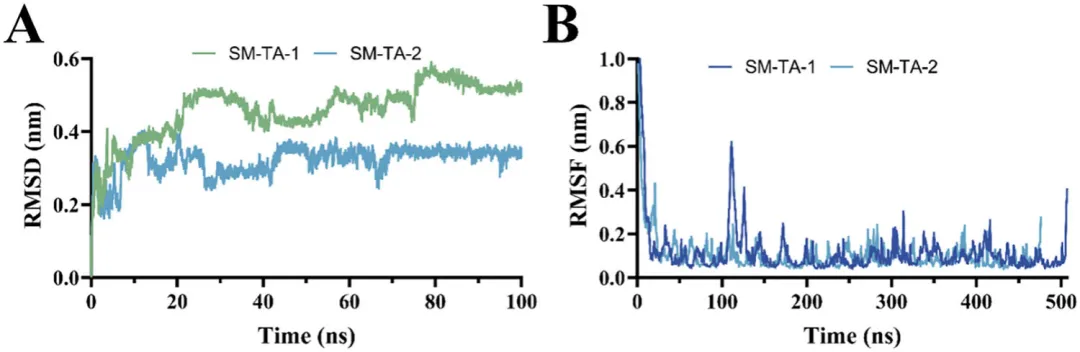
MD模拟是阐明蛋白质结构与稳定性之间关系的强大工具,其结果通常能与实验数据相互印证。通过计算模拟系统中主链RMSD随时间的变化,评估了SM-1-TA和SM-2-TA在MD模拟过程中的整体稳定性/平衡状态。RMSD是量化蛋白质构象波动的关键指标,较低的RMSD值表示较高的热稳定性。对SM-1-TA和SM-2-TA分别进行了100 ns的MD模拟,如图7A和7B所示。在整个模拟过程中,SM-2-TA系统迅速达到平衡,并保持稳定、低RMSD且波动极小。相比之下,SM-1-TA系统表现出更大且更持久的构象调整。因此,选择两个系统在75至100 ns的MD轨迹作为平衡轨迹进行后续分析。在模拟过程中,SM-2-TA的RMSD值始终高于SM-1-TA,表明其相对于初始结构有更大的结构偏差和构象变化。这也与SM-1-TA和SM-2-TA的最适温度和40°C温度稳定性的实验结果相符。同时,这也表明与SM-1-TA相比,SM-2-TA具有更大的应用潜力。
为了进一步研究构象动力学,计算了Cα原子的RMSF。如图7C和7D所示,虽然SM-1-TA和SM-2-TA在某些区域的RMSF谱图相似,但SM-2-TA在大部分结构中都表现出较低的波动幅度。这一结果与观察到的SM-2-TA更高的整体稳定性(较低的RMSD)相符。值得注意的是,SM-1-TA的无规则卷曲区域(残基100-150)显示出比SM-2-TA相应区域显著更高的RMSF值。这些发现表明,SM-1-TA比SM-2-TA具有更大的整体构象柔性,同时也表明该区域可能是影响SM-1-TA热稳定性的关键区域。在未来的研究中,针对该区域进行结构工程改造可能是增强SM-1-TA热稳定性的一种可行策略。
最后,为了阐明SM-2-TA比SM-1-TA具有更优异有机溶剂耐受性的潜在机制,分析了两种酶的溶剂可及表面积。SASA是分子可被溶剂分子接触的表面积,反映了其溶剂暴露程度。SM-2-TA表现出显著低于SM-1-TA的SASA值,表明其空间构象更紧凑。这种减小的SASA可能限制了与有机溶剂分子的相互作用,从而增强了其耐受性。
总之,MD模拟证实了SM-1-TA和SM-2-TA之间密切的结构-稳定性相关性。SM-2-TA表现出更稳定的构象、更低的结构柔性以及更紧凑的分子结构,这降低了其SASA,从而赋予其增强的热稳定性和有机溶剂耐受性。这些优越的结构和功能特性凸显了SM-2-TA作为有前景的工业应用生物催化剂的巨大潜力。
本研究为细菌单宁酶同工酶的生物技术开发和应用奠定了坚实的理论基础,并为未来的研究指明了方向。在未来的工作中,建议重点对SM-1-TA进行定点突变,以进一步提高其热稳定性。此外,通过工程化融合SM-1-TA和SM-2-TA构建融合蛋白,有可能产生具有更高催化效率和更强环境耐受性的新型单宁酶变体。Ba²⁺对SM-1-TA的激活作用也值得进一步研究其机制,以优化催化反应体系。同时,应进行大规模实验以验证SM-2-TA在食品脱酸、皮革废水生物修复和饲料生物加工等领域的实际工业适用性。此外,从链霉菌属物种中探索新的单宁酶同工酶,并阐明其功能互补的分子基础,将为微生物单宁酶资源的理性设计和生物学探索提供新策略,从而促进绿色生物催化产业的可持续发展。
本研究对来自阿维链霉菌的两种单宁酶同工酶SM-1-TA和SM-2-TA进行了系统的异源表达、酶学表征和结构-功能分析。值得注意的是,SM-1-TA和SM-2-TA均具有α/β水解酶家族的保守序列“Gly-X-Ser-X-Gly”及其催化三联体,这些在催化水解过程中起决定性作用。并且SM-1-TA和SM-2-TA在宽pH范围内对单宁底物表现出高效降解能力。其中,SM-2-TA表现出更高的热稳定性和对有机试剂的耐受性,而SM-1-TA对没食子酸甲酯和没食子酸丙酯表现出更高的催化效率。分子对接和动力学模拟进一步揭示了两种同工酶在结构柔性、底物结合模式和热稳定性方面的差异。SM-1-TA和SM-2-TA之间的功能互补性增强了宿主对环境波动的适应性,突显了同工酶在微生物系统中的生态学意义,同时为同工酶的进化和功能重要性提供了新的见解。
曾欢,食品科学硕士在读。以第一作者发表SCI论文3篇,主要研究方向为酶工程。
张鹏,南昌大学食品学院副教授,博士,硕士生导师。主要从事食品发酵、酶生物技术、合成生物学相关领域研究。入选江西省杰出青年基金计划项目,江西省科技特派员、江西省营养学会妇幼营养分会委员、南昌大学学报(理科版)青年编委、JAFC、JSFA等杂志审稿专家。近年来主持参与国家自然基金项目3项,江西省自然基金项目3项,江西省农业支撑项目1项、企业横向项目10项。在Journal of Agricultural and Food Chemistry、Food Hydrocolloids等国内外权威学术期刊上发表论文20余篇;参编英文专著1部,中文专著1部。
邓泽元,二级教授,博士生导师。南昌大学食品科学与资源挖掘全国重点实验室固定研究人员。享受国务院特殊津贴,赣鄱英才555工程人才、江西省新世纪百千万人才工程人选,江西省高校中青年学科带头人,首届江西省优秀硕士指导教师,江西省高等学校教学名师,获得教育部优秀中青年教师(支助),江西省主要学科学术与技术带头人,“食品科学”与“营养与食品卫生学”重点学科带头人。现任中国农学会微量元素与食物链分会副理事长,中国粮油学会营养分会副会长,中国营养学会营养转化医学分会副主任委员,江西省微量元素与健康研究会名誉理事长,江西省营养学会理事长;美国化学学会(ACS)会员、国际食品学会(IFT)会员,《食品科学》、《农产品加工学刊》、《食品安全质量检测学报》、《乳业科学与技术》、《南昌大学学报》(理科版)、J of Agri. Food Chem.和J of Food Biochem.编委,中国保健食品评审专家。