南昌大学殷军艺教授团队在Trends in Food Science & Technology发表重要综述
- 2026-06-01 22:41:31

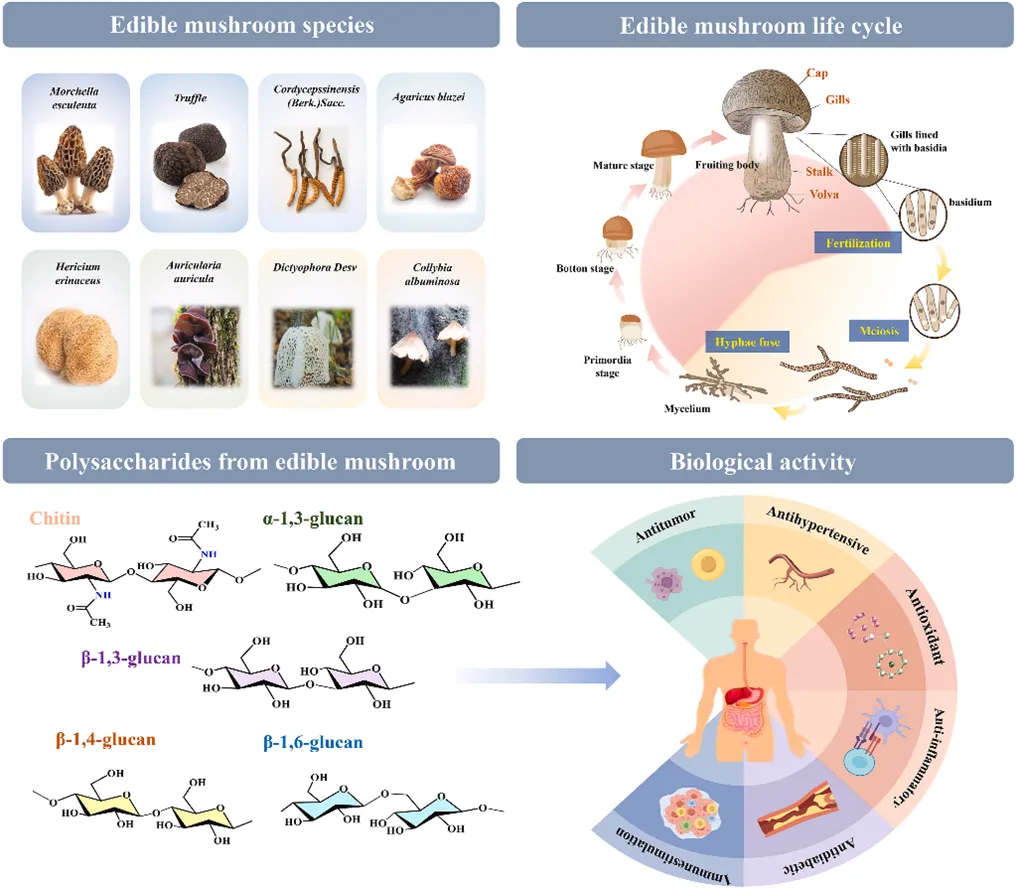
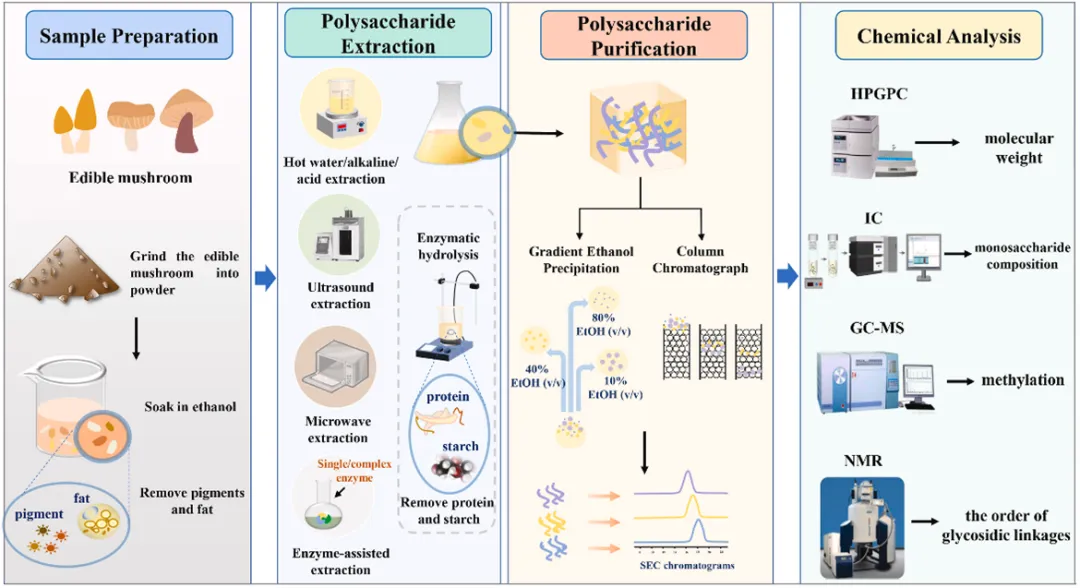
食用菌细胞壁组成
食用菌中碳水化合物的主要包括几丁质、葡聚糖和糖原。几丁质是一种由N-乙酰氨基葡萄糖通过β-1,4糖苷键聚合而成的线性多糖,为生物体提供结构支撑和保护。β-葡聚糖主要富集在食用菌细胞壁的最外层。糖原作为高度分支的葡聚糖,在食用菌中可能与β-葡聚糖共价连接而存在于细胞壁中。
食用菌细胞壁结构
真菌细胞壁的结构完整性高度依赖于特定多糖的精细结构。大多数真菌细胞壁结构具有层次结构。其最内层主要由链间葡聚糖和几丁质组成,形成一种动态的微纤维状结构,为细胞提供了一个核心网状支架,以承受质膜和细胞质施加的渗透压。细胞壁的外层由多糖、蛋白质及其他表面成分组成,这些成分共同构成了坚固且灵活的结构,使真菌能够适应其生理功能。细胞壁的完整性不仅取决于其组成,还受氢键和堆叠相互作用的影响。
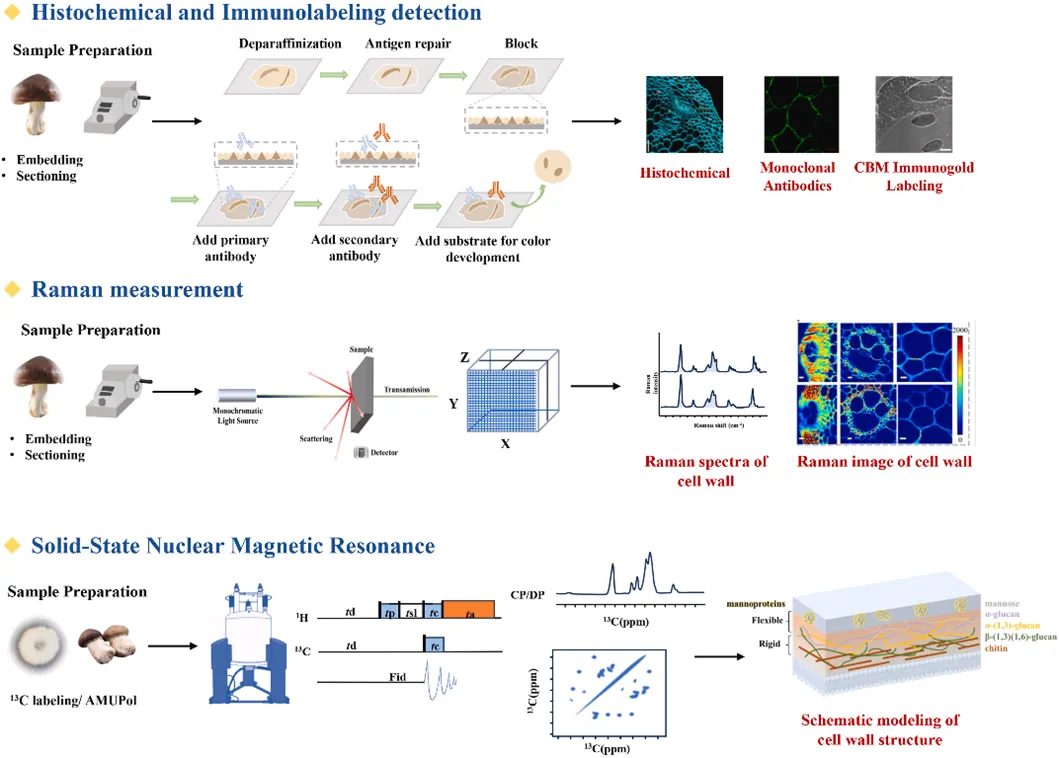
细胞壁多糖标记成像
细胞壁的可视化可为食用菌的形态学和细胞学观察提供重要的见解。基于细胞壁碳水化合物的化学特异性,组织化学和细胞化学方法通过特定染料与细胞壁中的不同成分发生反应,产生可见的颜色变化或荧光信号,如刚果红、钙荧光白、甲苯胺蓝和苯胺蓝染色法等。使用单克隆抗体(MAbs)和碳水化合物结合模块(CBMs)进行免疫荧光标记,可以在组织水平上对细胞壁多糖进行特异性识别和定位。
细胞壁多糖无标记成像
新的无标记成像技术已发展成为检测和表征细胞壁复杂化学和结构组成的强大工具。原子力显微镜(AFM)、傅立叶变换红外光谱(FTIR)、共聚焦拉曼显微镜(CRM)等技术可以提供细胞壁多糖的详细化学信息,有助于了解食用真菌细胞壁的复杂结构和功能。原子力显微镜与光谱学相结合可以获得细胞壁的化学成分,并阐明细胞壁的纳米结构、成分和功能之间的关系。拉曼光谱技术可对大型真菌中的脂质、蛋白质和多糖进行定量分析,为真菌学研究提供了新的基础数据。
多维固体核磁共振
多维固体核磁共振(ssNMR)作为一种前景广阔、功能强大的技术备受关注。与液体核磁共振不同,ssNMR 可在细胞或组织保持自然状态时进行测量,从而提供有关生物大分子的多态结构、分子间相互作用、空间排列、水接触和分子运动的详细数据。通过固体核磁共振技术结合高效液相色谱和气相色谱-质谱,提出了一种食用菌细胞壁模型。刚性核心由α-和β-葡聚糖、几丁质和岩藻糖组成。细胞壁的流动部分由β-葡聚糖和甘露糖组成。
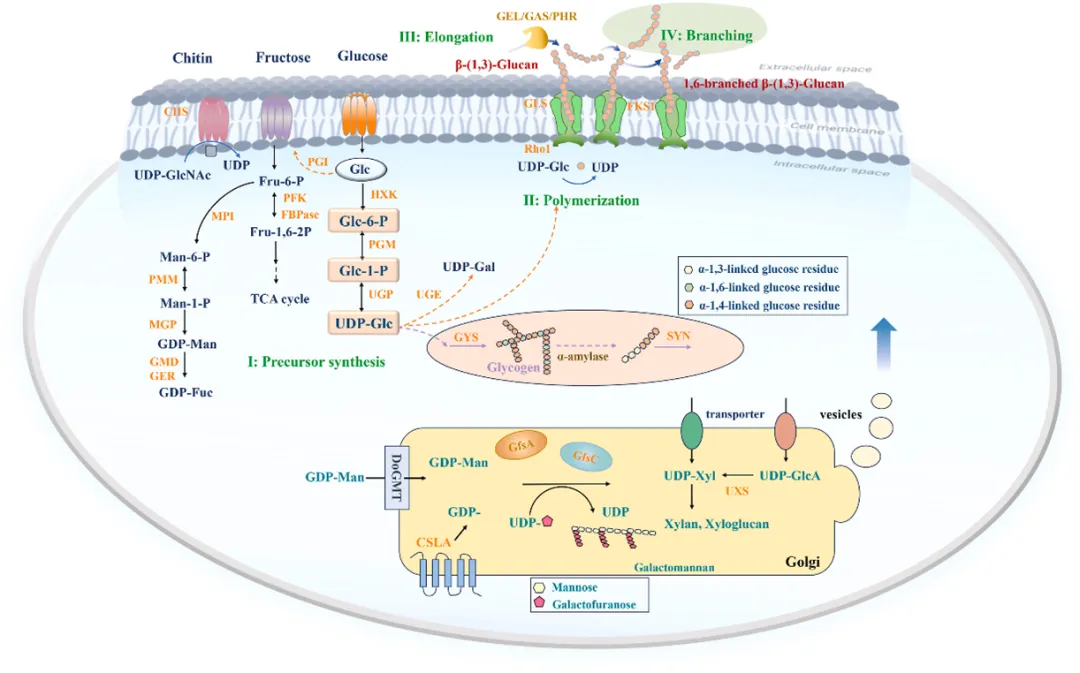
多糖的合成途径
糖链的形成大致可分为四个步骤:前体合成、短链聚合、长链延伸以及侧链分支。在反应过程中,蔗糖经历一系列转化生成UDP-糖,随后单糖通过不同的糖基转移酶从糖核苷酸供体转移到正在生长的多糖聚合物上。真菌中的短链线性β-1,3-葡聚糖由β-1,3-葡聚糖合成酶(GLS)生物合成。β-葡聚糖链在质膜上合成,随后在跨膜酶复合体的协助下转移至周质空间。被转移至质膜外层的长链β-1,3-葡聚糖,经多种细胞壁酶修饰后形成β-1,3-/1,6-支链。随后,β-1,3-葡聚糖经多种糖基转移酶的作用发生延伸、分支并与其他组分交联,从而形成细胞壁典型的复杂多糖网络。
多糖合成中的关键酶
多糖的合成需要不同酶的协同作用,这些酶可根据其功能分为核苷酸-糖合成酶、聚合酶、分支酶、异构酶和转移酶。通过核苷糖合成酶(如 UGP、UGDG、UGE)调节前体供应与单糖多样性,聚合酶与分支酶控制主链延伸及分支结构以影响物理化学性质,以及异构酶(如 PGM、PGI、PMM)定向引导碳源流向,从分子水平决定了多糖的精细结构与生物活性。糖基转移酶(GTs)是多糖生物合成中的核心转移酶,通过高尔基室的层级定位和对囊泡运输的动态调控催化特定糖苷键的形成,从而决定EMP的精细结构和生物功能。
多糖的顺序合成与平衡合成
高尔基体通过其特有的糖基转移酶和功能分区(cis、medial、trans),以顺序合成或复合体介导的模式精准调控多糖的侧链分支与修饰,与在质膜合成的葡聚糖显著不同;同时,食用菌通过由转录因子、碳通量代谢及细胞壁完整性(CWI)信号通路构成的多级调节网络,感应环境压力并调控基因表达,从而在不同生长阶段维持多糖合成的动力学平衡与结构稳定性。这一平衡合成机制的解析,为深入理解食用菌多糖的构效关系提供了关键的理论支撑。
(请点击标题查看)
分子对接与分子动力学黄酮类化合物
食品感官科学与分析农产品贮藏与加工
肉与肉制品蛋与蛋制品水产品
奶及奶制品豆及豆制品果蔬及果蔬制品
大米及米制品
炎症性肠病糖尿病肝病神经疾病
(请点击标题查看)
(请点击学者姓名查看)


在线投稿平台链接:https://www.wjx.top/jq/85151447.aspx
标注‘原创’仅代表原创编译,本平台不主张对原文的版权。本平台转载仅仅是出于学术交流和传播信息的需要,并不意味着代表本平台观点或证实其内容的真实性;转载文章版权归原作者所有,作者如果不希望被转载或有侵权行为,请联系本平台删除。由于编译水平有限,推文或简历有不妥之处,深感抱歉,请联系本平台修改或者删除。

