南昌大学第二附属医院 | 揭示年龄相关性听力损失新机制:耳蜗内质网应激介导 Cx26 泛素化降解!
- 2026-05-25 15:30:33

概述
以年龄相关性听力损失(ARHL)中耳蜗连接蛋白 26(Cx26)降解机制不明为切入点,先通过 C57BL/6J 小鼠模型验证年龄增长与耳蜗 Cx26 下调、内质网(ER)应激标志物上调的相关性,再经体外细胞、耳蜗外植体实验及体内药物诱导 ER 应激模型,探究 ER 应激对 Cx26 表达的影响,最后通过转录组分析、免疫荧光共定位等明确分子机制,得出结论:耳蜗 ER 应激通过激活 Nedd4 和 Eps15 介导的泛素 - 蛋白酶体通路,加速 Cx26 降解及缝隙连接斑解体,这是 ARHL 的重要致病机制,为靶向 ER 应激或泛素化调控的治疗提供新方向。
注:该文章发表于《iScience》,该期刊最新影响因子为4.1,位列JCR Q1区。
研究要点解析
本研究首次明确耳蜗内质网(ER)应激与年龄相关性听力损失(ARHL)中连接蛋白 26(Cx26)降解的因果关系,揭示 Nedd4 和 Eps15 介导的泛素 - 蛋白酶体通路是核心机制。通过体内外多模型验证,发现 ER 应激激活早于听力损失出现,且 Cx26 降解是 ARHL 早期病理事件,为 ARHL 的早期干预提供了新靶点,同时提出调控耳蜗蛋白稳态(靶向 ER 应激或泛素化通路)的治疗新范式,突破了现有 ARHL 缺乏有效药物治疗的困境。
动物模型:采用 C57BL/6J 小鼠(1 月龄、6 月龄、12 月龄)作为 ARHL 模型,通过腹腔注射衣霉素(TCM)构建 ER 应激体内模型;
细胞与外植体实验:用 BXPC3 细胞,经二硫苏糖醇(DTT)、TCM 诱导 ER 应激,同时培养耳蜗外植体,结合蛋白酶体抑制剂 MG132 验证降解途径;
检测技术:采用听觉脑干反应(ABR)评估听力阈值,通过 RT-qPCR 检测 mRNA 表达,免疫荧光染色观察蛋白定位与缝隙连接斑结构,Western blot 定量蛋白水平,转录组分析筛选泛素化相关基因,免疫荧光共定位验证蛋白相互作用。
小鼠耳蜗 Cx26 呈年龄依赖性下调,12 月龄小鼠 Cx26 蛋白水平较 1 月龄下降 89.4%,缝隙连接斑碎片化且长度缩短,而 Gjb2 mRNA 水平稳定;
老年小鼠耳蜗 ER 应激标志物(GRP78、PDI、GRP94、CHOP)的 mRNA 和蛋白水平均显著上调;
体外实验中,DTT、TCM 可浓度依赖性降低 BXPC3 细胞和耳蜗外植体的 Cx26 蛋白水平,MG132 可逆转该降解;
TCM 诱导的 ER 应激小鼠模型中,耳蜗 Cx26 蛋白减少、缝隙连接斑解体,但短期内未出现明显听力损失和毛细胞丢失;
ER 应激显著上调耳蜗中 Nedd4、Eps15 等泛素化相关基因表达,且 Nedd4、Eps15 与 Cx26 在细胞 membrane 共定位。
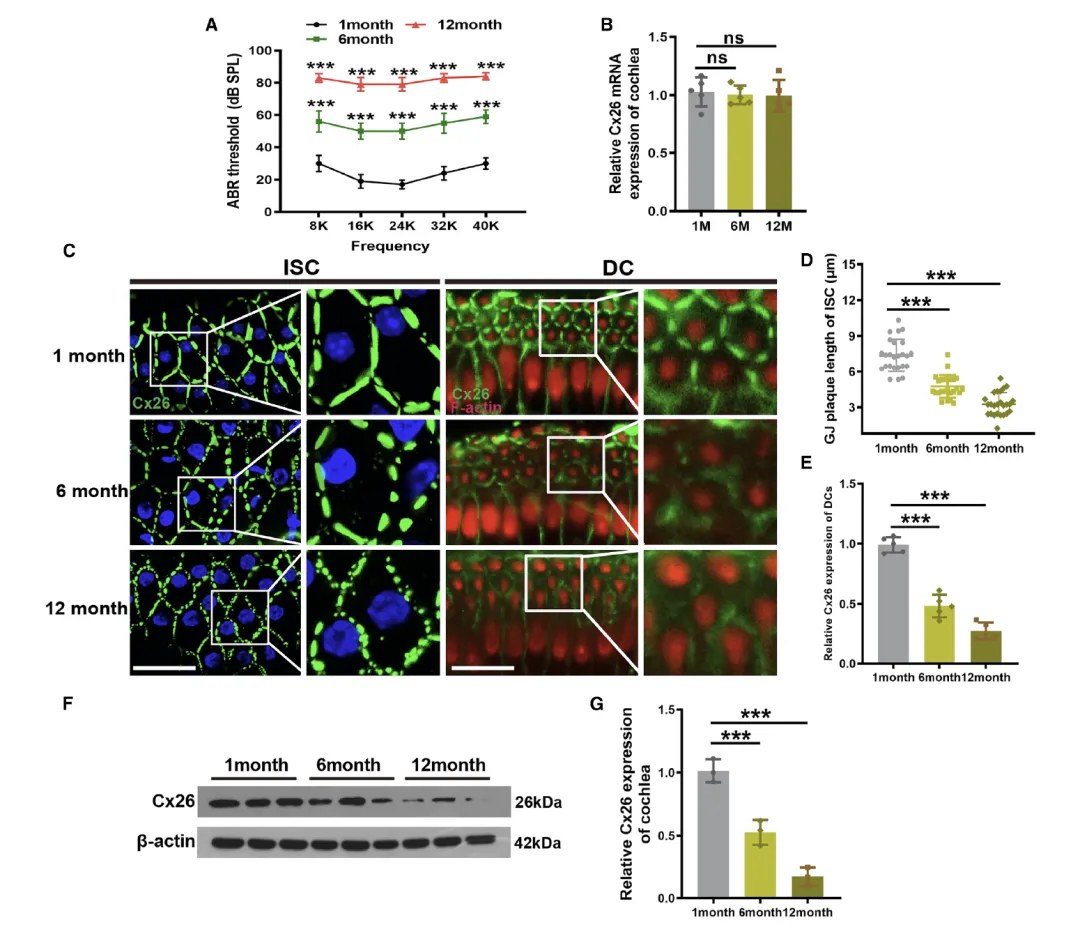
Figure 1:小鼠耳蜗 Cx26 表达的年龄依赖性下降及缝隙连接斑(GJP)结构重塑
该图通过多维度实验验证了 C57BL/6J 小鼠耳蜗中 Cx26 随年龄增长的动态变化与听力功能衰退的关联。A 图的听觉脑干反应(ABR)测试显示,与 1 月龄小鼠相比,6 月龄小鼠的听力阈值显著升高,12 月龄时进一步恶化,明确了该小鼠模型符合年龄相关性听力损失(ARHL)的病理特征。B 图的 RT-qPCR 结果表明,不同年龄组小鼠的 Gjb2 mRNA 水平无显著差异,提示 Cx26 的表达调控发生在转录后层面。C 图的免疫荧光染色直观呈现了内沟细胞(ISCs)中 GJP 的结构变化:1 月龄小鼠的 Cx26 呈特征性线性排列,6 月龄时出现离散碎片化,12 月龄时仅残留散在的点状结构。D 图对 ISCs 中连续 GJP 长度的定量分析,以及 E 图对 Deiter 细胞(DCs)中 Cx26 荧光强度的检测,均证实了 Cx26 的表达量和 GJP 完整性随年龄增长呈渐进性下降。F 和 G 图的 Western blot 结果进一步量化了这种变化,6 月龄和 12 月龄小鼠的 Cx26 蛋白水平分别较 1 月龄下降 50.7±7.2% 和 89.4±5.8%,综合证明了年龄相关的 Cx26 蛋白降解和 GJP 解体是 ARHL 的重要病理表现。
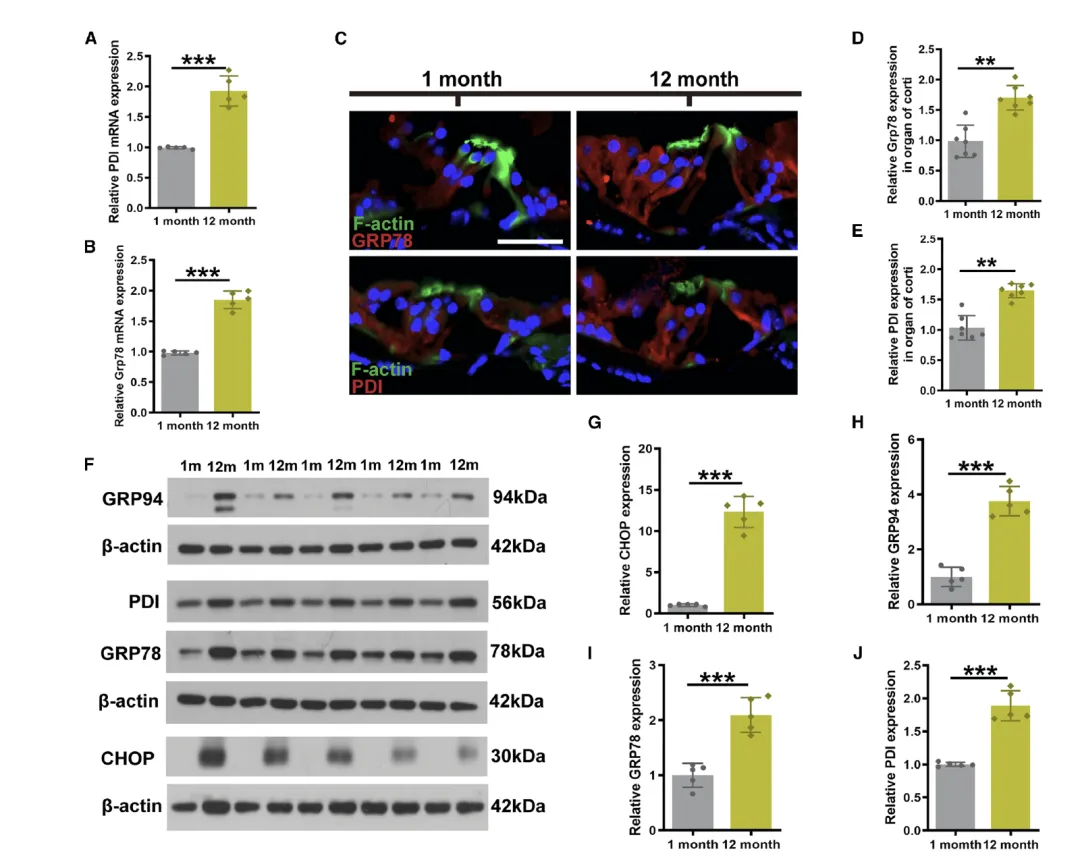
Figure 2:老年 C57BL/6J 小鼠耳蜗内质网(ER)应激信号通路激活
该图聚焦于老年小鼠耳蜗中 ER 应激的激活状态,为 ER 应激与 ARHL 的关联提供了直接证据。A 和 B 图的 RT-qPCR 结果显示,与 1 月龄年轻小鼠相比,12 月龄老年小鼠耳蜗中 ER 应激标志物葡萄糖调节蛋白 78(GRP78)和蛋白二硫键异构酶(PDI)的 mRNA 表达水平显著上调,表明 ER 应激在转录层面被激活。C 图的免疫荧光染色进一步定位了这些标志物的表达部位,发现 GRP78 和 PDI 的荧光信号在老年小鼠的柯蒂氏器中明显增强,证实 ER 应激主要发生在耳蜗的关键功能区域。D 和 E 图对荧光强度的定量分析,客观验证了老年小鼠耳蜗中 GRP78 和 PDI 的表达量显著高于年轻小鼠。F 图的 Western blot 结果则从蛋白水平拓展了这一发现,除 GRP78 和 PDI 外,另两种 ER 应激相关蛋白 GRP94(分子伴侣)和 CHOP(促凋亡效应因子)的表达也随年龄增长显著升高,G-J 图的定量分析进一步确认了这四种蛋白的年龄依赖性上调,表明老年小鼠耳蜗中未折叠蛋白反应(UPR)被显著激活,ER 应激通路处于异常活跃状态。
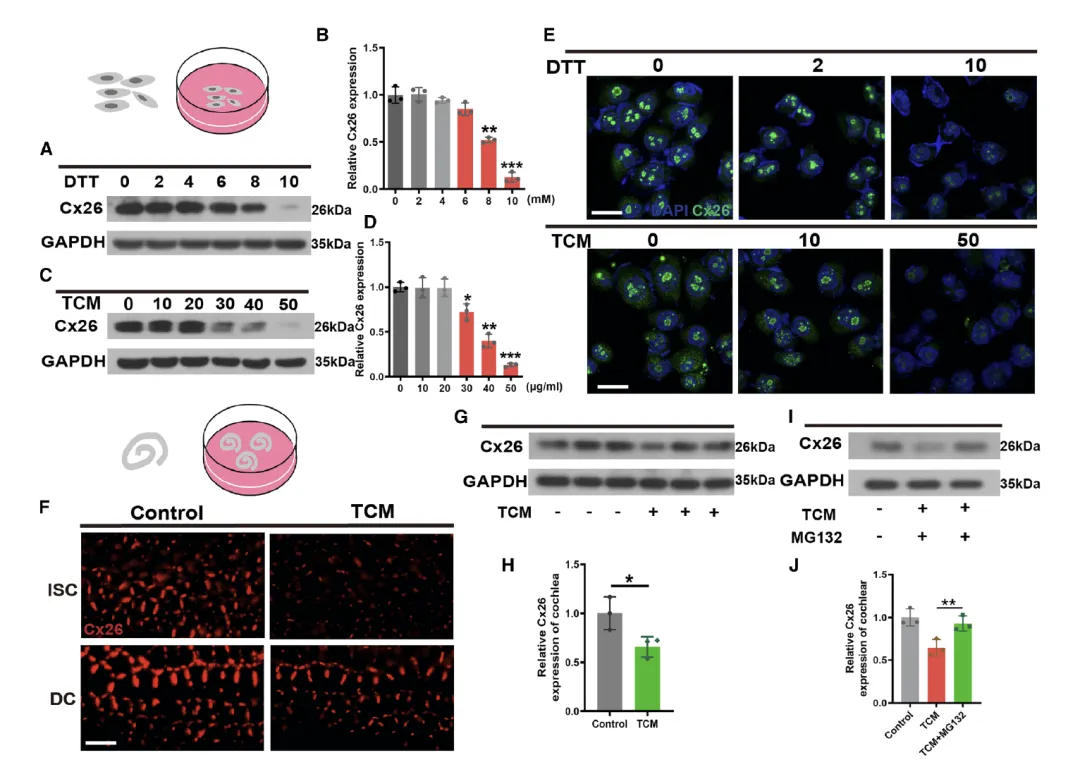
Figure 3:内质网(ER)应激在体外模型中降低 Cx26 表达
该图通过细胞和耳蜗外植体的体外实验,直接证实 ER 应激对 Cx26 表达的抑制作用及相关降解途径。A 和 B 图显示,用 ER 应激诱导剂二硫苏糖醇(DTT)处理 BXPC3 细胞后,Cx26 蛋白表达呈浓度依赖性下降,8 mM 和 10 mM DTT 处理 6 小时后,Cx26 表达显著降低,表明 DTT 介导的 ER 应激可直接影响 Cx26 水平。C 和 D 图则用另一种 ER 应激诱导剂衣霉素(TCM)得到类似结果,浓度低至 30 μg/mL 的 TCM 即可有效抑制 Cx26 表达,进一步验证了 ER 应激与 Cx26 下调的因果关系。E 图的免疫荧光染色直观显示,低浓度 DTT(2 mM)和 TCM(10 μg/mL)对 Cx26 表达无明显影响,而高浓度(10 mM DTT、50 μg/mL TCM)则显著降低 Cx26 的荧光信号。F 图对耳蜗外植体的免疫荧光染色表明,TCM 处理后,DCs 和 ISCs 中的 Cx26 免疫反应性明显减弱;G 和 H 图的 Western blot 定量结果确认了外植体中 Cx26 蛋白水平的降低,提示 ER 应激介导的 Cx26 降解可能通过蛋白酶体途径实现。I 和 J 图进一步验证了这一机制,蛋白酶体抑制剂 MG132 与 TCM 联合处理后,Cx26 的降解被显著抑制,直接证明 ER 应激通过蛋白酶体途径促进 Cx26 降解。
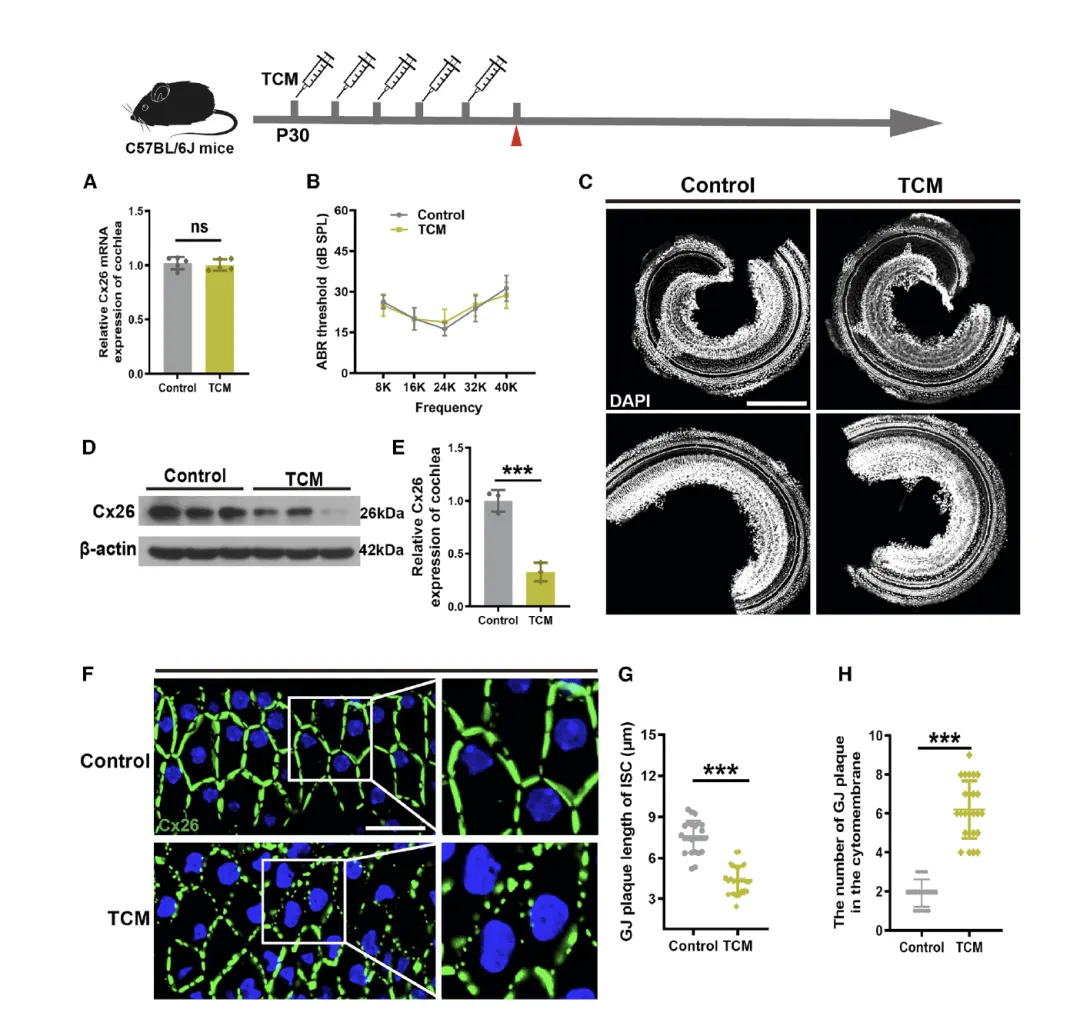
Figure 4:体内内质网(ER)应激降低 C57BL/6J 小鼠耳蜗 Cx26 表达并破坏缝隙连接斑(GJP)结构
该图通过体内药物诱导模型,证实 ER 应激在活体小鼠耳蜗中对 Cx26 的调控作用,且这种调控先于听力损失发生。A 图的 RT-qPCR 结果显示,经 TCM 处理的小鼠与对照组相比,Cx26 的 mRNA 水平无显著差异,与体外实验一致,表明 ER 应激对 Cx26 的调控发生在转录后水平。B 图的 ABR 测试结果显示,连续 TCM 处理未导致小鼠听力阈值显著升高,C 图的免疫荧光染色也未发现柯蒂氏器毛细胞丢失,提示在该实验条件下,ER 应激诱导的 Cx26 变化先于明显的听觉功能损伤和细胞死亡。D 和 E 图的 Western blot 结果显示,TCM 处理组小鼠耳蜗的 Cx26 蛋白水平显著低于对照组,证实体内 ER 应激可有效降低 Cx26 蛋白表达。F 图的免疫荧光染色显示,对照组小鼠 ISCs 中的 Cx26 形成连续的线性 GJP 结构,而 TCM 处理组的 GJP 碎裂为不连续的点状结构;G 和 H 图的定量分析表明,TCM 处理组的 GJP 长度显著缩短,斑块数量显著增加,说明 ER 应激不仅降低 Cx26 表达,还破坏其形成的缝隙连接结构,导致耳蜗细胞间通讯能力受损。
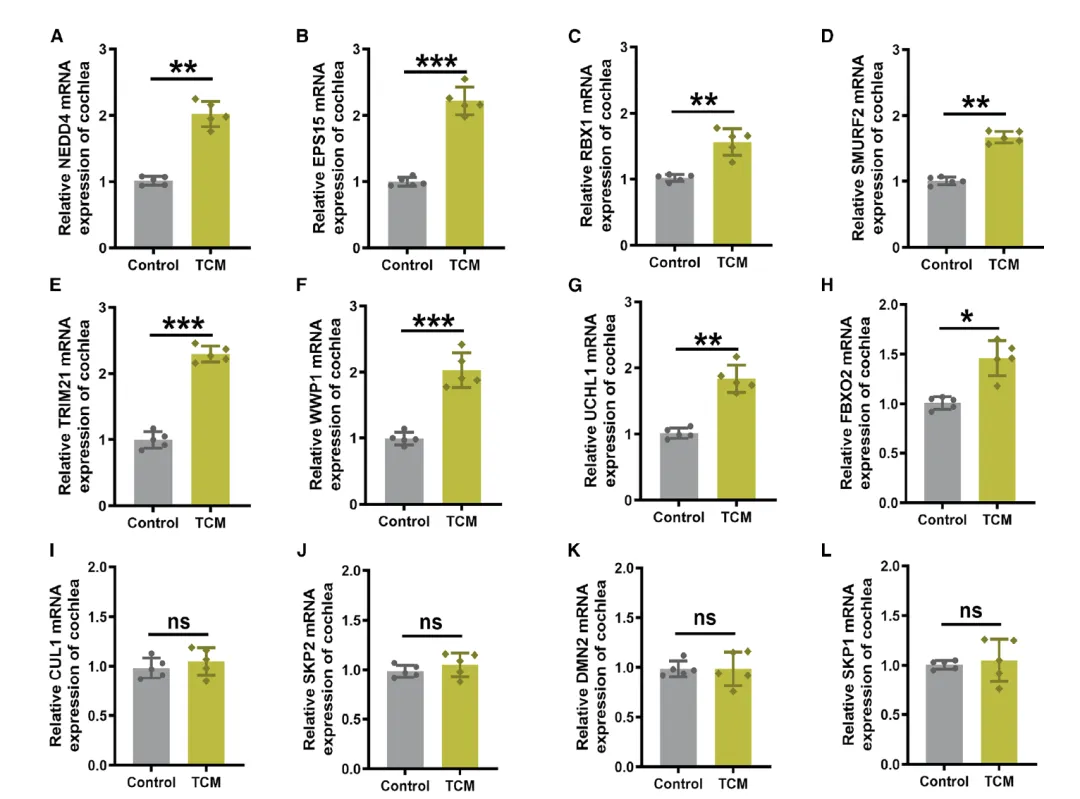
Figure 5:内质网(ER)应激上调耳蜗中泛素化相关基因的 mRNA 表达
该图通过转录组分析,揭示 ER 应激激活泛素化相关基因表达,为 Cx26 降解的分子机制提供线索。A-H 图的 RT-qPCR 结果显示,TCM 诱导 ER 应激后,小鼠耳蜗中一系列泛素化相关酶和蛋白降解相关基因的转录水平显著上调,包括泛素连接酶 NEDD4、衔接蛋白 EPS15,以及 RBX1、SMURF2、TRIM21、WWP1、UCHL1、FBXO2 等,这些基因均参与泛素 - 蛋白酶体通路的调控,提示该通路在 ER 应激介导的 Cx26 降解中可能发挥关键作用。I-L 图则显示,另一部分泛素化相关基因如 CUL1、SKP2、DNM2、SKP1 的 mRNA 水平在对照组和 TCM 处理组之间无显著差异,表明 ER 应激对泛素化通路的激活具有选择性,并非全面上调所有相关基因,而是针对性激活特定的泛素化调控因子,为后续明确 Cx26 降解的关键介导分子提供了方向。
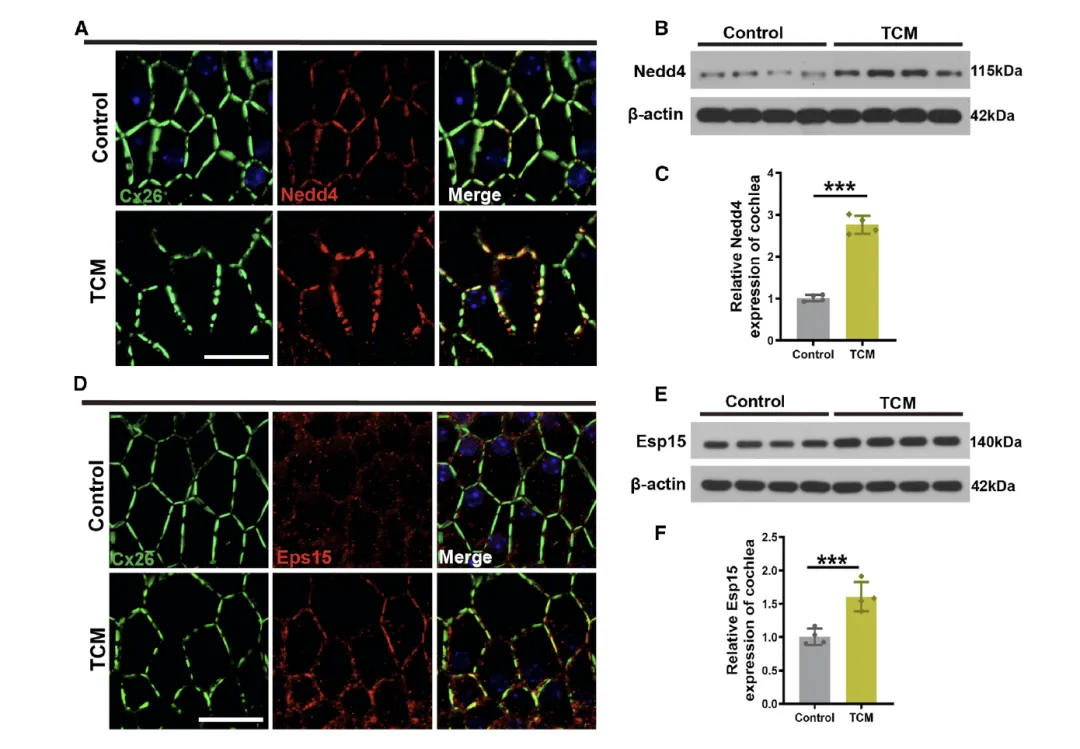
Figure 6:内质网(ER)应激增加耳蜗中泛素化相关蛋白 Nedd4 和 Eps15 的表达并与 Cx26 共定位
该图通过蛋白水平检测和共定位分析,明确 Nedd4 和 Eps15 是 ER 应激介导 Cx26 泛素化降解的关键分子。A 图的免疫荧光共定位结果显示,对照组小鼠 ISCs 中,Nedd4(红色)与 Cx26(绿色)在细胞膜上存在共定位,而 TCM 处理组中,ISCs 的 GJP 结构严重碎裂,细胞膜上的 Nedd4 表达显著增强,且与碎片化的 Cx26 共定位更明显,提示 Nedd4 可能直接参与 Cx26 的调控。B 和 C 图的 Western blot 结果证实,TCM 处理组小鼠耳蜗中 Nedd4 的总蛋白水平显著高于对照组,进一步验证了 ER 应激对 Nedd4 的上调作用。D 图的免疫荧光共定位显示,TCM 处理后,ISCs 中 Eps15(红色)的表达显著增加,且与碎片化的 Cx26(绿色)大量共定位,表明 Eps15 可能与 Nedd4 协同参与 Cx26 的降解过程。E 和 F 图的 Western blot 定量结果确认了 TCM 处理组中 Eps15 蛋白水平的显著升高,综合证明 ER 应激通过上调 Nedd4 和 Eps15 的表达,促进其与 Cx26 的相互作用,进而介导 Cx26 的泛素化降解。
4.结论
本研究证实,耳蜗内质网(ER)应激是年龄相关性听力损失(ARHL)中连接蛋白 26(Cx26)降解的主要原因。ER 应激通过激活泛素 - 蛋白酶体通路,促进 Nedd4 和 Eps15 介导的 Cx26 泛素化,加速其降解并破坏缝隙连接斑完整性,且这一过程早于听力损失和毛细胞损伤发生。该发现首次揭示了 ER 应激与 Cx26 降解的分子关联,明确了 ARHL 的新致病机制,为开发靶向 ER 应激通路或泛素化调控因子(如 Nedd4、Eps15)的药物提供了科学依据,有望延缓 ARHL 进展。
局限性:未探究 Cx26 在耳蜗血管纹中间细胞和螺旋韧带中的年龄相关变化,未明确 Cx26 缝隙连接斑破坏对通道功能的直接影响,且未证实 Cx26 降解与毛细胞退化的因果关系。展望:未来需采用更合适的动物模型和先进检测技术,深入研究上述未明确的问题,同时探索抗氧化与调控 ER 应激结合的干预策略,为 ARHL 的临床治疗提供新方案。
文章来自:Bai X, Liao B, Hu WH, Wang MQ, Sun Y, Chen XB, Xu K. Cochlear endoplasmic reticulum stress causes connexin 26 degradation is involved in age-related hearing loss. iScience. 2025 Oct 28;28(12):113891. doi: 10.1016/j.isci.2025.113891. PMID: 41321625; PMCID: PMC12663731.
#年龄相关性听力损失#耳蜗#连接蛋白263Cx26#内质网应激#泛素化#Nedd4#Eps15#缝隙连接
本文中使用的图片来源于Pubmed,因客观原因未能与权利人取得联系。本平台出于学术交流目的引用,无意侵犯原作者权益。如权利人认为不妥,请及时联系公众号后台,我们将立即删除或协商解决。

国家杰青一对一答疑视频
医学省自然申请答疑,立项的关键条件是哪一些?从哪些方向可以杀出重围
临床型博士如何准备国青标书?没有预实验怎么办?专家一对一解答规划
中医药科研研究