一、从四色到二色:哺乳动物的“降级”
3亿年前,脊椎动物祖先拥有四色视觉:四种视锥蛋白——LWS(红色)、SWS1(紫色/紫外)、SWS2(蓝色)、RH2(绿色)——共同工作,今天的鸟类和爬行动物仍保留这套“全彩系统”。进入恐龙时代(三叠纪晚期),早期哺乳动物为适应夜行生活,首先丢失了RH2(绿色);之后在有袋类和真兽类(胎盘类)的演化中进一步丢失了SWS2(蓝色),最终退化为只剩LWS(红/长波)和SWS1(紫外/短波)的二色视觉——这正是狗、猫等大多数哺乳动物是红绿色盲的根本原因。
二、灵长类的“逆袭”:三色视觉的诞生
约6500万年前恐龙灭绝后,灵长类祖先重返白天,以果实和嫩叶为食。分辨红色成熟果实与绿色未熟果实的生存压力,驱动了自然选择。三色视觉为灵长类带来了巨大的演化优势,其中最主流的解释是“觅食假说”:拥有三色视觉的个体能更高效地在茂密的绿叶中发现色彩鲜艳的成熟果实。研究表明,三色视觉的个体在检测红色或橙色食物方面比二色视觉的个体更具优势——对恒河猴的长期观察发现,拥有三色视觉的雌性在定位和取食果实方面速度明显更快。事实上,三色视觉的演化与果实颜色的演化密切相关,两者在漫长的岁月中形成了一场双向塑造的“协同进化”:果树进化出鲜艳的果皮颜色来吸引灵长类帮助传播种子,而灵长类则进化出更敏锐的色觉来获取营养丰富的食物。这一相互选择的过程,正是达尔文自然选择理论的绝佳体现。
新大陆猴(中南美洲的猴子)和旧大陆猴/猿类(人类的祖先,人类属于旧世界灵长类分支)在约3500万年前分道扬镳。新大陆猴的X染色体上没有发生基因复制,而是靠同一个LWS基因位点上的多个“版本”(等位基因)实现色觉差异——只有携带两个不同版本的雌猴才能看到三色,雄性永远只有二色。有趣的是,这些不同版本与旧大陆猴复制出的L和M基因序列极为相似,表明早在两类猴子分化之前,L基因的不同光谱版本就已存在;而旧大陆猴和猿类的祖先后来通过基因复制,把这些原本分散在不同个体中的版本固定成了两个独立的基因,从而获得了稳定的三色视觉——这便是人类三色视觉的进化起源。
三、为什么红绿色盲男性更多?
L和M视蛋白的基因均定位于X染色体,二者紧密连锁、序列同源性极高。男性仅携带一条X染色体,若该染色体上的L或M基因发生突变或重组异常,会直接引发红绿色盲;女性拥有两条 X 染色体,当其中一条携带缺陷基因时,另一条正常染色体可代偿其感光功能。因此,男性红绿色盲率约7%,女性仅约0.5%——这正是红绿色盲成为人类最常见色觉缺陷的原因。
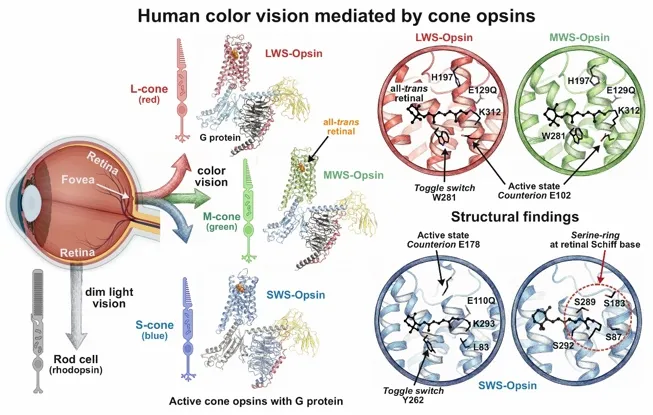
四、大脑的“调色板”:三种视锥蛋白如何调配出百万种颜色?
人类感知颜色,并非眼睛直接“看到”颜色,而是对光的物理属性进行神经转化。物体反射不同波长的光进入眼睛后,被视网膜上的三种视锥细胞接收。大脑就像一位画家,根据三种细胞激活比例调配出相应的颜色——比例1:0:0是红色,1:1:0是黄色,1:1:1则是白色。
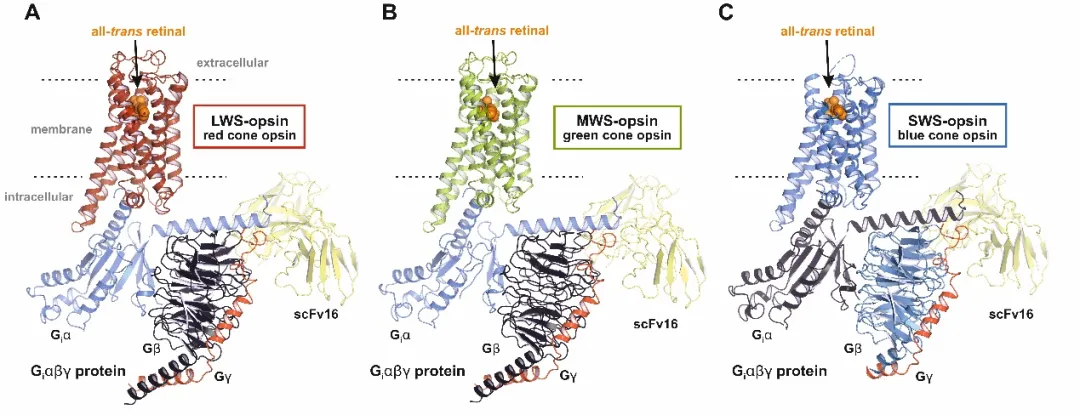
然而,L与M视蛋白序列相似度高达96%,为何一个感红、一个感绿?S视蛋白与它们差异显著,又如何感知蓝光?视锥细胞为何比视杆细胞反应快得多?尽管视杆蛋白(Rhodopsin)的结构早在2000年就已被解析(这也是首个被解析的GPCR结构),但视锥蛋白的高分辨率三维结构却长期缺失。
本研究利用冷冻电镜技术,首次解析了三种人视锥蛋白——红敏(LWS-opsin)、绿敏(MWS-opsin)、蓝敏(SWS-opsin)——的活性态高分辨率结构,为我们揭开了这幅“调色板”背后的分子机制。