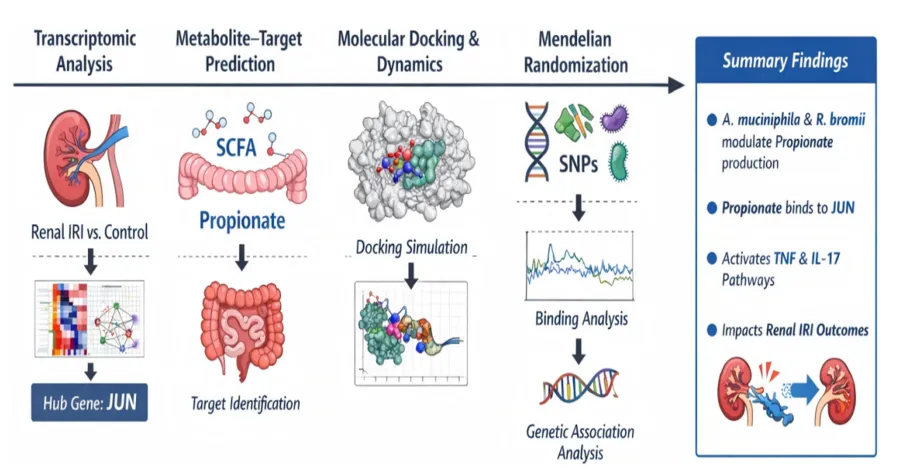
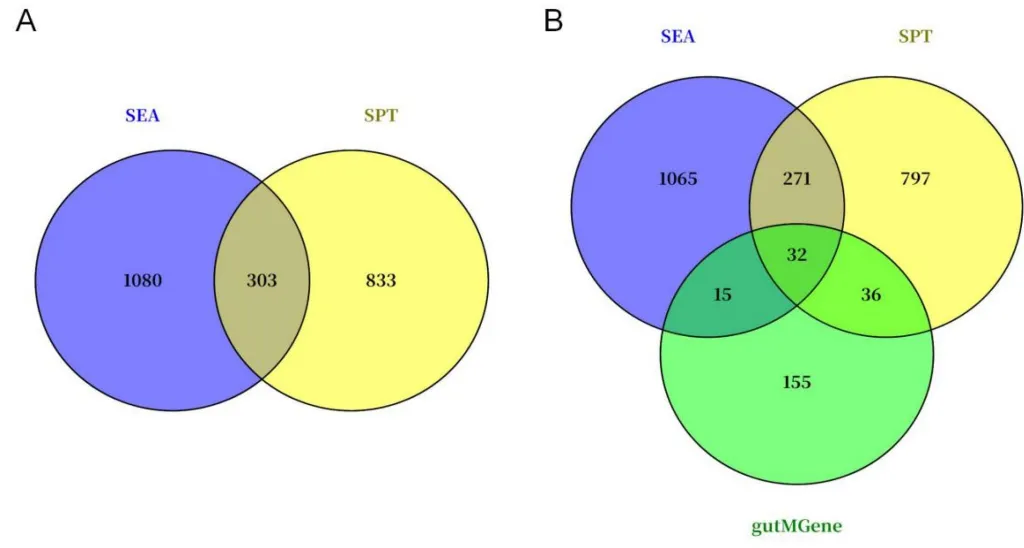
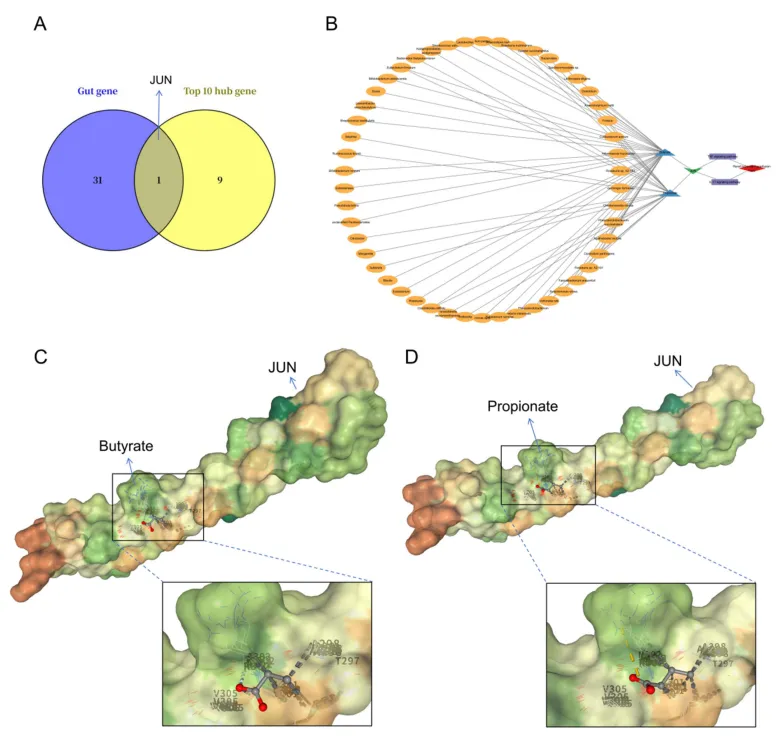
从gutMGene数据库获取277种菌群代谢物和238个人类肠道靶基因,经SEA与STP数据库分别预测出1383和1136个潜在靶点。取两数据库交集得到303个共同靶点,再与gutMGene收录的靶基因重叠,最终筛选出32个肠道微生物代谢物的核心人类靶基因。这一精简的靶点集合覆盖了短链脂肪酸等关键代谢物的作用分子,为后续将微生物代谢信号与肾IRI转录组数据对接提供了精准的候选基因池,其中JUN随后被识别为唯一重叠的关键交汇分子(图2)。
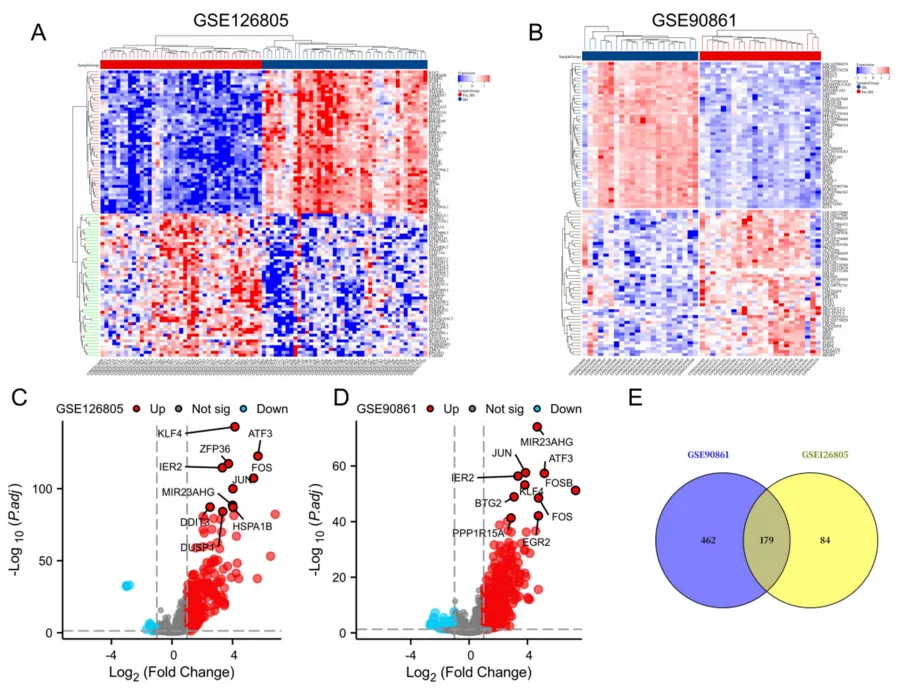
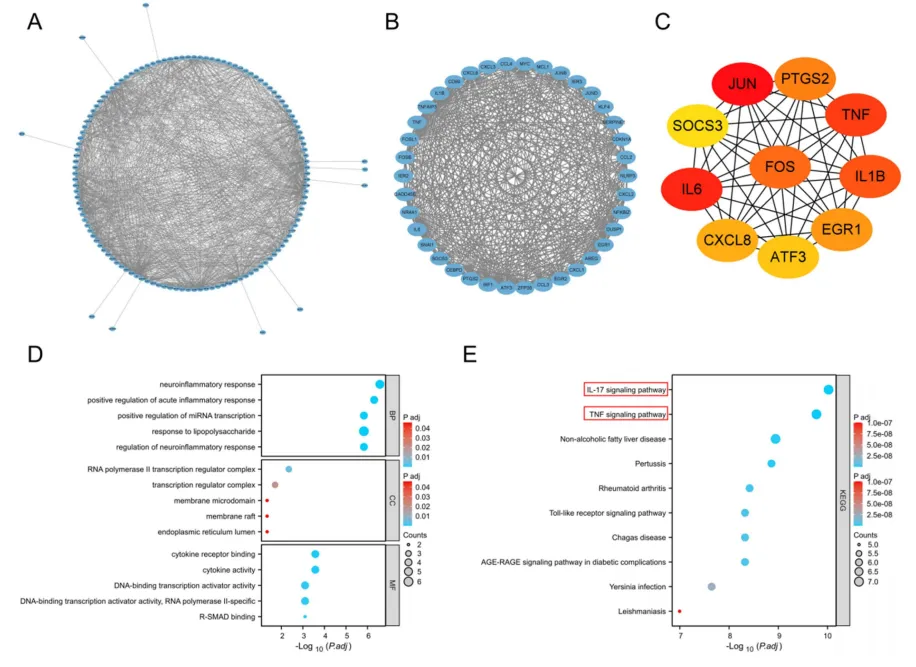
分析GSE126805和GSE90861两个肾IRI转录组数据集,分别鉴定出263个和641个差异表达基因(|log2FC|>1,调整p<0.05)。热图与火山图均显示IRI组与对照组的表达谱显著分离,JUN、FOS、ATF3等即刻早期基因普遍上调。取两数据集交集获得179个共同差异基因,这些基因在IRI中呈现一致性的转录改变,主要富集于炎症反应与氧化应激相关通路,为后续构建蛋白互作网络并识别核心调控因子提供了可靠的分子基础(图3)。
图3:肾缺血再灌注损伤表达谱鉴定的共有差异表达基因
基于179个共同差异基因构建PPI网络,利用MCODE提取高连接模块,再通过cytoHubba的MCC算法排序,获得JUN、FOS、TNF、IL6、PTGS2等10个hub基因。GO富集分析显示这些基因参与神经炎症反应及脂多糖应答,KEGG通路则显著富集于IL-17和TNF信号轴。该结果揭示了JUN等核心基因紧密交织于经典炎症通路之中,提示它们可能作为上游代谢信号向下游效应分子传递的关键转录调控节点,在肾IRI中发挥枢纽作用(图4)。
维恩图显示JUN是唯一同时属于32个代谢物靶点与10个IRI hub基因的分子。构建的“菌群-代谢物-靶点-通路-疾病”网络将丁酸盐和丙酸盐连接到JUN,进而指向IL-17/TNF通路。分子对接显示丁酸盐与丙酸盐对JUN蛋白的结合能分别为-13.4kcal/mol和-12.1kcal/mol,均形成稳定氢键。这表明JUN在肠道微生物代谢物与肾脏炎症之间扮演结构生物学桥梁,短链脂肪酸可通过直接结合JUN蛋白来潜在调控其转录活性(图5)。
图5:JUN作为肠道来源调控靶点可受短链脂肪酸结合调控
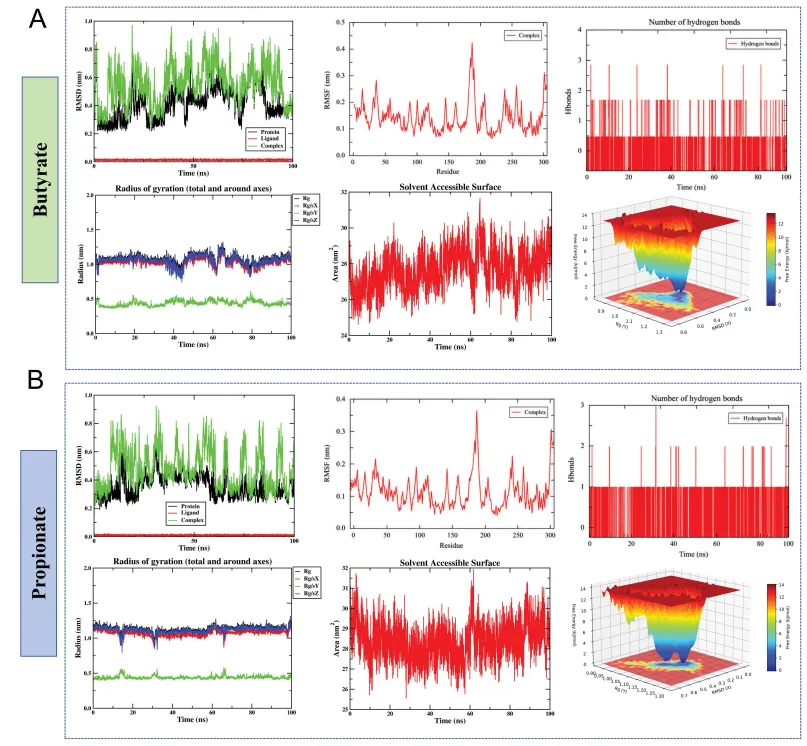
对丁酸盐-JUN和丙酸盐-JUN复合物进行100 ns分子动力学模拟。丙酸盐体系的骨架RMSD更快达到平衡,Rg和SASA波动更小,自由能景观呈现单一集中的低能盆地;而丁酸盐体系则显示稍高的构象灵活性和多个亚稳态。氢键分析表明两者均为弱且瞬时的相互作用。综合轨迹参数,丙酸盐与JUN的结合稳定性和构象收敛性均优于丁酸盐,提示丙酸盐可能是更适配的生理配体,指导后续研究聚焦丙酸盐(图6)。
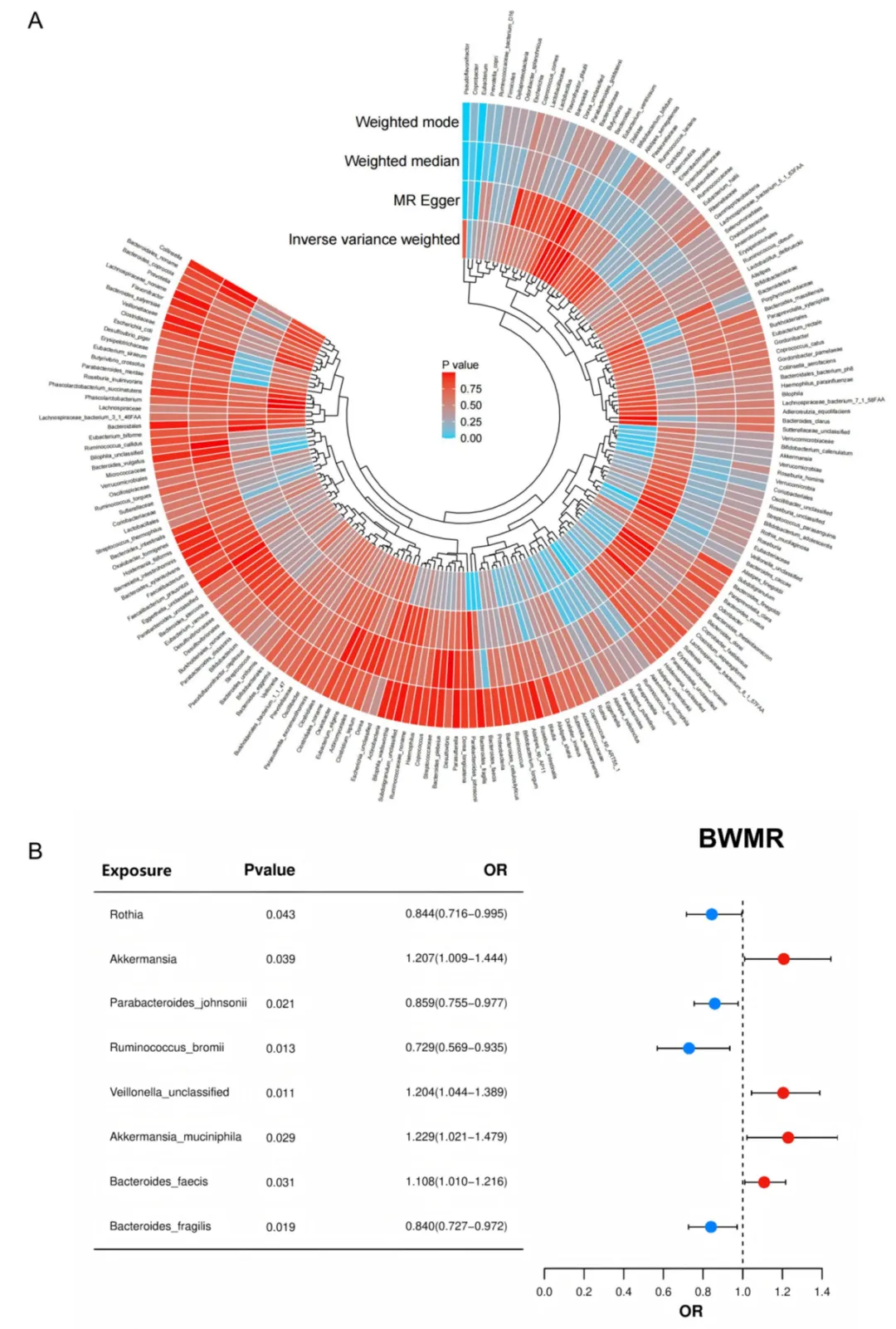
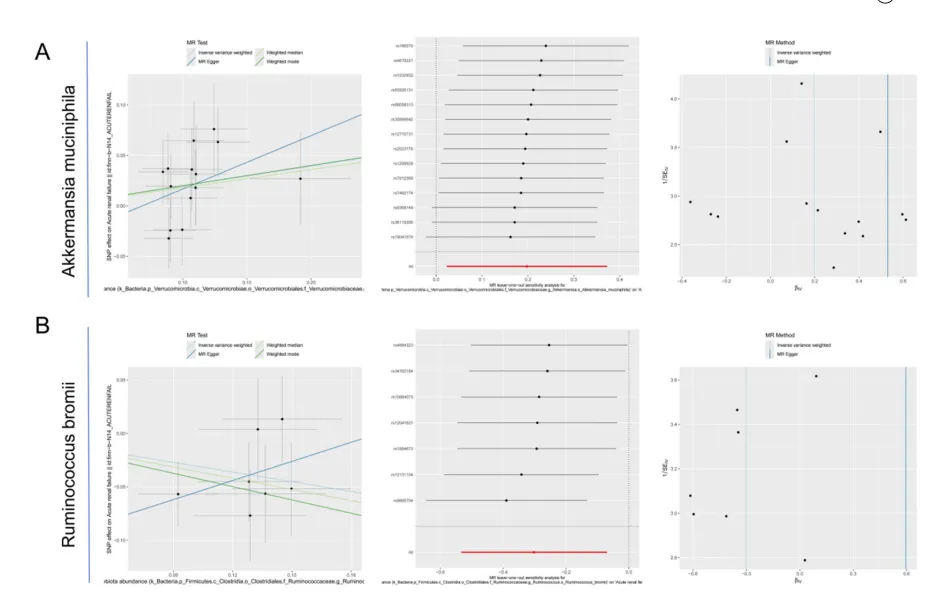
以207个肠道菌群为暴露、急性肾损伤为结局进行双样本孟德尔随机化分析。逆方差加权法结合贝叶斯加权孟德尔随机化验证,鉴定出8个与肾损伤易感性显著相关的菌群。其中Akkermansia muciniphila呈正向因果关联(OR=1.219,p=0.026),Ruminococcus bromii呈负向保护关联(OR=0.740,p=0.011)。两个关键菌种均参与丙酸盐代谢,为上游“菌群-丙酸盐-JUN”调控轴提供了来自遗传学的因果证据支持(图7)。
针对Akkermansia muciniphila和Ruminococcus bromii分别绘制散点图、森林图和漏斗图。各单核苷酸多态性效应方向一致,MR-Egger截距检验未检测到水平多效性(p>0.05),Cochran’s Q检验亦未发现显著异质性(p>0.05)。反向孟德尔随机化分析未显示反向因果关联。这些敏感性分析证实两个菌群与肾损伤易感性之间的因果估计稳健可靠,不受混杂多效性干扰,进一步强化了二者作为潜在干预靶点的可信度(图8)。